
 |
 |
 |
 |
 |
Artigo
|
|
| Discriminação de madeiras similares por fluorescência molecular e mínimos quadrados parciais Discrimination between similar woods by molecular fluorescence and partial least squares |
|
Elian Meneses OliveiraI; Jez Willian Batista BragaII,*; Alexandre Florian da CostaI
IDepartamento de Engenharia Florestal, Faculdade de Tecnologia, Universidade de Brasília, 70919-970 Brasília - DF, Brasil Recebido em 05/05/2015 *e-mail: jez@unb.br Wood is an extremely complex biological material, which can show macroscopic similarities that make it difficult to discriminate between species. Discrimination between similar wood species can be achieved by either anatomic or instrumental methods, such as near infrared spectroscopy (NIR). Although different spectroscopy methods are currently available, few studies have applied them to discriminate between wood species. In this study, we applied a partial least squares-discriminant analysis (PLS-DA) model to evaluate the viability of using direct fluorescence measurements for discriminating between Eucalyptus grandis, Eucalyptus urograndis, and Cedrela odorata. The results show that molecular fluorescence is an efficient technique for discriminating between these visually similar wood species. With respect to calibration and the validation samples, we observed no misclassifications or outliers. INTRODUÇAO A madeira é um material biológico extremamente complexo, amplamente conhecido e utilizado. Porém, macroscopicamente, algumas espécies podem apresentar semelhanças, dificultando, assim, uma identificaçao confiável da espécie. A distinçao ou discriminaçao de madeiras pode ser feita tanto por critérios científicos, levando em conta características anatômicas e organolépticas, quanto por procedimentos ou práticas populares, considerando apenas as características organolépticas ou nomes comuns. A identificaçao anatômica de espécies madeireiras é um processo relativamente complexo, que requer treinamento especializado, além de uma coleçao de madeiras disponíveis para comparaçoes entre espécies, de acordo com o nível de detalhamento exigido.1 Processos nao-anatômicos, como o teste de fluorescência e a espectroscopia do infravermelho próximo (NIR) também podem auxiliar na identificaçao de diferentes espécies, sendo estas técnicas emergentes e que exigem a realizaçao de uma calibraçao para a obtençao de um método eficiente que permita a identificaçao de espécies produtoras de madeiras.1-4 A fluorescência é uma etapa do processo de desativaçao, ou seja, processo pelo qual uma molécula excitada volta ao seu estado fundamental, havendo emissao de um fóton de radiaçao. A trajetória favorecida para o estado fundamental é aquela que minimiza o tempo de vida do estado excitado. Assim, se a desativaçao por fluorescência é rápida e mais favorável em relaçao aos processos nao radiativos, tal emissao é observada.5 A técnica de espectroscopia de fluorescência tem sido uma ferramenta eficaz para a mediçao de pequenas quantidades de substâncias químicas, devido à sua alta sensibilidade e seletividade.6 O primeiro registro de observaçao do fenômeno fluorescência em madeiras se deu há mais de 400 anos, com a espécie de Lignum nephriticum. Esta espécie era bem conhecida na Europa, nos séculos XVI e XVII, devido aos copos ou taças que eram feitos desta madeira exótica, que exibia uma curiosa cor azul na superfície da água colocada no copo.7 A fluorescência em madeiras pode ser visualizada tanto por métodos simples que usam uma lanterna de ultravioleta (UV) para observar uma superfície recém-cortada da madeira (avaliaçao visual) ou por métodos mais elaborados, como o uso de espectrofluorímetros, que permite a obtençao de espectros. Estudos realizados expondo diferentes espécies de madeiras à luz UV indicaram que a fluorescência é uma importante característica de diagnóstico na identificaçao de madeira e distinçao entre cerne e alburno.8-11 Observa-se que, dependendo da espécie, as madeiras apresentam fluorescências diferentes, com a cor variando de marrom ao violeta com praticamente todos os tons de laranja, amarelo, verde, azul e anil. A intensidade da fluorescência, no entanto, nao está limitada a uma área particular da madeira. Em alguns casos, o cerne fluoresce de forma mais intensa que o alburno e, em outros casos ocorre o inverso.9 Por outro lado, em algumas espécies a fluorescência emitida quando a madeira é irradiada com luz UV é imperceptível a olho nu. Além disso, madeiras macroscopicamente semelhantes podem também apresentar fluorescência semelhante em aspectos de cor e intensidade da cor, dificultando o processo de discriminaçao entre espécies, sendo, entao, necessária a utilizaçao de uma técnica instrumental para a mediçao da fluorescência para fins de identificaçao correta da espécie produtora de madeira. Portanto, o objetivo deste estudo foi realizar medidas de fluorescência molecular emitida por madeiras de três espécies florestais, Eucalyptus grandis, Eucalyptus urograndis e Cedrela odorata, investigando o potencial desta técnica instrumental para fins de discriminaçao destas espécies, empregando calibraçao multivariada por mínimos quadrados parciais para análise discriminante (PLS-DA, do inglês Partial Least Squares for Discriminant Analysis).
PARTE EXPERIMENTAL O estudo foi conduzido no Laboratório de Automaçao, Quimiometria e Química Ambiental (AQQUA) do Instituto de Química da Universidade de Brasília, em colaboraçao com o Departamento de Engenharia Florestal da Faculdade de Tecnologia da Universidade de Brasília. Obtençao das amostras e preparo do material As amostras das espécies florestais Eucalyptus grandis, Eucalyptus urograndis e Cedrela odorata foram obtidas a partir de peças maiores cedidas pelo Laboratório de Produtos Florestais (LPF) do Serviço Florestal Brasileiro (SFB), pertencente ao Ministério do Meio Ambiente (MMA). Essas amostras foram identificadas por especialistas em anatomia de madeira do LPF/SFB e escolhidas por serem visualmente semelhantes entre si.12 Foram confeccionadas 90 amostras de dimensoes uniformes (2 x 2 x 2 cm), sendo 30 amostras de cada espécie. Inicialmente, as 90 amostras foram secas em estufa a 50 ºC, até massa constante. Em seguida, a superfície radial das amostras das madeiras foi lixada com lixas de papel gra nº 180 e 400, respectivamente, para retirada de camada oxidada, já que o processo de oxidaçao pode interferir na correta análise das amostras. Determinaçao da fluorescência Os espectros de emissao de fluorescência das espécies estudadas foram obtidos com o auxílio do aparelho espectrofluorímetro da marca Varian, modelo Cary Eclipse (Austrália), com acessório para medidas de amostras sólidas. Para a realizaçao das medidas foram utilizadas fendas de 5 nm em ambos os monocromadores de excitaçao e o de emissao, velocidade de varredura em 1200 nm/min, ganho da fotomultiplicadora em 600 V e com a radiaçao ultravioleta fornecida por uma lâmpada de xenônio pulsada de 50 W. Foram adquiridos espectros de emissao de fluorescência entre 380 e 660 nm com excitaçao nos comprimentos de onda de 370, 400, 430, 460, 490, 520 e 550 nm. Construçao dos modelos de discriminaçao Foram desenvolvidos três modelos, sendo que cada um discriminaria uma espécie em relaçao às outras. Para a construçao dos modelos de discriminaçao, os dados obtidos a partir das 90 amostras foram divididos aleatoriamente em dois conjuntos, sendo um usado para calibraçao (20 amostras por espécie) e o outro para validaçao (10 amostras por espécie). O modelo utilizado para a discriminaçao das espécies foi o PLS-DA, o qual já se encontra descrito em detalhes na literatura.13-16 O software utilizado para o desenvolvimento dos modelos de discriminaçao foi o PLS Toolboxr, versao 6.5, o qual é executado por meio do programa Matlabr, versao R2011a. Conforme descrito por Botelho et al., a validaçao de modelos de discriminaçao pode ser atestada pelo cálculo de figuras de mérito distintas das geralmente empregadas em modelos quantitativos, sendo estas: a taxa de erros falso positivo, taxa de erros falso negativo, a seletividade, a especificidade e a taxa de eficiência do método.13 Contudo, em casos onde têm-se uma discriminaçao clara entre as classes, sem ocorrência de erros de classificaçao, a observaçao apenas dessas figuras de mérito durante a otimizaçao do modelo nao permite a determinaçao das melhores condiçoes de pré-processamento, regiao espectral e número de variáveis latentes. Pois, nesse caso, diferentes condiçoes podem apresentar valores idênticos para todas as figuras de mérito mencionadas. Nessas situaçoes, a otimizaçao do modelo pode ser realizada observando os valores dos erros médios, expressos em termos da raiz quadrada do erro médio quadrático de calibraçao (RMSEC, do inglês root mean square error of calibration), validaçao cruzada (RMSECV, do inglês root mean square error of cross-validation) e a distribuiçao dos valores de classe estimados para as amostras de calibraçao. A princípio, as condiçoes que apresentarem o menor valor de RMSECV e que nao sobreajustarem o modelo, o que pode ser comprovado na validaçao pela comparaçao dos valores de RMSEC e raiz quadrada do erro médio quadrático de previsao (RMSEP, do inglês root mean square error of prediction), fornecerao o modelo com menor probabilidade de erros de classificaçao.
RESULTADOS E DISCUSSAO O espalhamento de radiaçao é um fenômeno comum e que pode ser detectado ao se realizar as medidas de fluorescência. Esses espalhamentos ocorrem quando a energia incidente em uma direçao é desviada para outras direçoes, havendo a produçao de radiaçao difusa.17 Observou-se na Figura 1 que o espalhamento presente nas medidas é conhecido como Rayleigh, ocorrendo no mesmo comprimento de onda de excitaçao e, por este motivo, se desloca de acordo com a mudança do comprimento de onda de excitaçao. Esses espalhamentos atrapalham a modelagem, devendo ser excluídos do espectro para a construçao dos modelos de discriminaçao. Desvios de linha de base também sao observados nos espectros de uma mesma espécie. Para corrigir tais desvios, visando obter menores erros de discriminaçao, foi escolhido o pré-processamento "Variável Normal Padrao" (SNV, do inglês Standard Normal Variate).
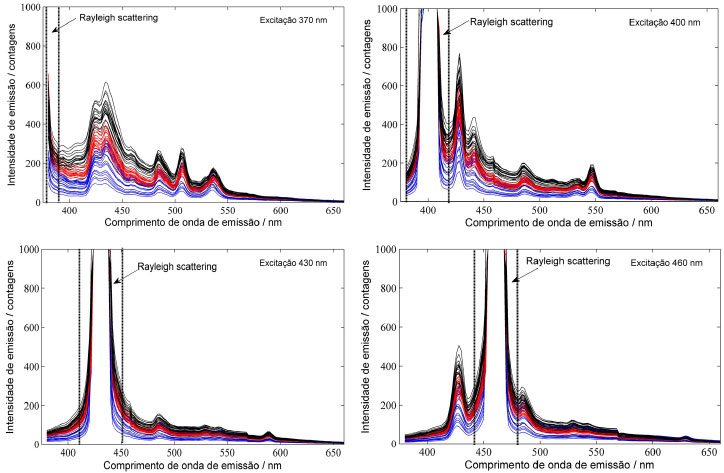 Figura 1. Espectros de emissao de fluorescência obtidos em diferentes comprimentos de onda de excitaçao. Cedrela odorata (azul); Eucalyptus grandis (vermelho) e Eucalyptus urograndis (preto)
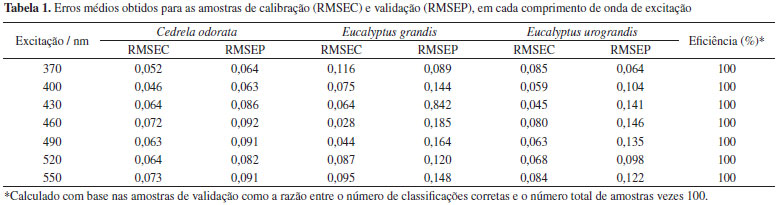
Além das características já mencionadas, também pode ser observado na Figura 1 que as três espécies apresentam espectros muito semelhantes, com as mesmas bandas de emissao. Contudo, observa-se também que há uma diferença de intensidade de emissao entre as espécies. A espécie Cedrela odorata apresenta menor intensidade do sinal de fluorescência, seguida por Eucalyptus grandis e Eucalyptus urograndis. Este comportamento facilita a discriminaçao entre as espécies. Entretanto, os espectros ainda apresentam intensidades muito próximas, dificultando a análise por inspeçao visual. Além disso, a variaçao presente dentro de uma mesma espécie faz com que haja uma sobreposiçao entre os espectros de espécies distintas. Portanto, para se obter uma discriminaçao com maior segurança, evitando erros na identificaçao da espécie, é necessária uma modelagem dos espectros a partir de um modelo de discriminaçao multivariado. Para o método apresentado nesse trabalho, foi observada uma clara discriminaçao das classes por meio da modelagem por PLS-DA, de forma que os valores de RMSEC, RMSECV e a distribuiçao dos valores de classe estimados foram utilizados para a otimizaçao do método. A Tabela 1 apresenta os valores de RMSEC, RMSEP e a taxa de eficiência em percentagem para os modelos construídos nos comprimentos de onda de excitaçao em 370 nm, 400 nm, 430 nm, 460 nm, 490 nm, 520 nm e 550 nm. Em todos os modelos a taxa de eficiência obtida foi de 100%. Portanto, o melhor comprimento de onda de excitaçao a ser utilizado para a discriminaçao das espécies pode ser determinado observando os resultados dos erros médios (RMSEC e RMSEP), sendo que os menores valores desses parâmetros e que nao apresentem diferença estatisticamente significativa entre si indicaram o melhor o comprimento de excitaçao a ser escolhido. Erros com estas características implicam na ausência de sobreajuste do modelo, ou seja, o erro médio obtido para as amostras utilizadas para a calibraçao nao apresentará uma diferença significativa do erro médio obtido para as amostras de validaçao do modelo. Portanto, pelo critério dos erros médios, o comprimento de onda de excitaçao escolhido foi 370 nm. Esse resultado é concordante com o fato dos espectros de emissao com excitaçao em 370 nm apresentarem um número maior de bandas de emissao e maior intensidade.
Partindo do modelo desenvolvido com os espectros de emissao obtidos com o comprimento de onda de excitaçao em 370 nm, os gráficos de dispersao dos valores de classe obtidos para a discriminaçao das três espécies (Cedrela odorata, Eucalyptus grandis e Eucalyptus urograndis) sao apresentados nas Figuras 2, 3 e 4, respectivamente.
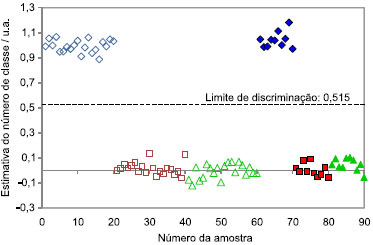 Figura 2. Dispersao das amostras de calibraçao (símbolos vazios) e validaçao (símbolos cheios) para discriminaçao da espécie Cedrela odorata outras duas espécies. (  ) Cedrela odorata, ( ) Cedrela odorata, ( ) Eucalyptus grandis e ( ) Eucalyptus grandis e ( ) Eucalyptus urograndis ) Eucalyptus urograndis
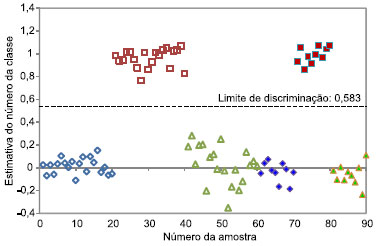 Figura 3. Dispersao das amostras de calibraçao (símbolos vazios) e validaçao (símbolos cheios) para discriminaçao da espécie Eucalyptus grandis outras duas espécies. ( ) Cedrela odorata, () Eucalyptus grandis e () Eucalyptus urograndis
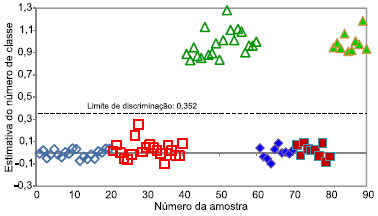 Figura 4. Dispersao das amostras de calibraçao (símbolos vazios) e validaçao (símbolos cheios) para discriminaçao da espécie Eucalyptus urograndis outras duas espécies. ( ) Cedrela odorata, () Eucalyptus grandis e () Eucalyptus urograndis
O limite de discriminaçao representa o valor que melhor separa as duas classes. Caso uma amostra apresente valor de classe superior ao limite de discriminaçao, esta amostra é identificada como pertencente à espécie que o modelo está discriminando, enquanto que, se o valor de classe for inferior ao limite de discriminaçao, esta amostra é pertencente às demais espécies. Este limite de discriminaçao é determinado com base na dispersao das amostras de calibraçao, de forma a minimizar a ocorrência de erros de classificaçao. Caso as dispersoes das duas classes sejam muito semelhantes, o limite de discriminaçao fica próximo a 0,5, valor que corresponde ao centro da escala. Caso contrário, há um deslocamento do limite de discriminaçao, visando minimizar possíveis erros de classificaçao. Este comportamento pode ser notado nas Figuras 2, 3 e 4, em que o limite de discriminaçao é 0,515, 0,583 e 0,352, para Cedrela odorata, Eucalyptus grandis e Eucalyptus urograndis, respectivamente. Para a discriminaçao da espécie Eucalyptus urograndis pode-se observar que as amostras de calibraçao dessa espécie apresentam uma variaçao superior às das outras duas espécies, provocando o deslocamento do limite de discriminaçao para 0,352. Ao analisar as Figuras 2, 3 e 4, observa-se que a espécie discriminada possui estimativas de valores de classe próximos a 1, diferente das outras duas espécies, que apresentam valores de classe próximos a 0, evidenciando que nao existe erro sistemático presente nos modelos. Além disso, comparando a discriminaçao da espécie Cedrela odorata (cedro) em relaçao às demais espécies, observa-se uma menor dispersao dos valores de classe estimados, evidenciando um melhor ajuste do modelo de discriminaçao para esta espécie. Este resultado é concordante com o fato dessa espécie ser a mais distinta entre as três estudadas. Além disso, a dispersao das amostras de calibraçao é bem semelhante à dispersao das amostras de validaçao, confirmando assim, a ausência de sobreajuste do modelo. Amostras anômalas ("outliers") sao definidas como amostras que apresentam um erro de estimativa do número de classe relativamente maior que a maioria das amostras de calibraçao.18 Observando a Figura 2 (Cedrela odorata), verifica-se visualmente que nenhuma amostra pode ser considerada anômala. Entretanto, nas Figuras 3 e 4 (Eucalyptus grandis e Eucalyptus urograndis, respectivamente), sao observadas algumas amostras de calibraçao que poderiam ser identificadas como anômalas (amostras número 28 para Eucalyptus grandis e número 52 para Eucalyptus urograndis), devido a um maior distanciamento das demais amostras pertencentes à mesma espécie. Entretanto, devido ao número reduzido de amostras do conjunto de dados e da ausência de erros de classificaçao, decidiu-se por nao excluir nenhuma amostra. Observando-se ainda as Figuras 2, 3 e 4, verifica-se que as três espécies foram discriminadas corretamente, uma vez que os gráficos de dispersao confirmam os baixos erros citados anteriormente e uma clara separaçao entre as classes foi observada. Ademais, nenhuma amostra, de calibraçao e de validaçao, se aproxima do limite de discriminaçao, mesmo na presença de possíveis amostras anômalas, demonstrando que o método apresenta uma margem de confiança considerável para a identificaçao das espécies, principalmente para Cedrela odorata (cedro), caso em que as amostras estao mais distantes do limite de discriminaçao. Para os modelos finais nao foram observados erros falso positivo ou falso negativo, os valores de sensibilidade e especificidade obtidos foram iguais aos seus valores máximos (1,0) e a eficiência igual a 100%.
CONCLUSAO As medidas de fluorescência molecular aliadas a modelos PLS-DA possibilitaram a obtençao de um método eficiente para discriminar espécies de madeira macroscopicamente semelhantes, uma vez que os gráficos de dispersao apresentaram-se bastante claros em relaçao à distinçao entre a espécie discriminada e as demais espécies em estudo. Apesar da semelhança entre os espectros de emissao, o melhor comprimento de onda para discriminar corretamente as espécies foi de 370 nm. A dispersao dos dados foi menor para Cedrela odorata (cedro), mostrando um melhor ajuste do modelo de discriminaçao para esta espécie. Pelo nosso conhecimento, nao foram encontrados trabalhos empregando a técnica de fluorescência molecular para a discriminaçao de madeiras. Contudo, comparando os erros médios e as dispersoes obtidas nesse trabalho com os resultados publicados empregando espectrômetros NIR de bancada para discriminar madeiras, pode-se concluir que as duas técnicas possuem eficiência similar.3,4 Apesar dos ótimos resultados obtidos pelo método proposto, é observado que a fluorescência em madeiras pode ser influenciada por diversos fatores ambientais, tais como a temperatura, exposiçao da amostra à chuva, presença de umidade, etc. Portanto, novos estudos vêm sendo realizados em nosso grupo de pesquisa para avaliar a influência desses fatores e na discriminaçao de um número maior de espécies, além de verificar a potencialidade desta técnica para aplicaçao em campo para fins de fiscalizaçao do transporte ou comercializaçao de madeiras.
AGRADECIMENTOS Os autores agradecem a o CNPq (processo 307223/2012-4), ao Laboratório de Produtos Florestais do Serviço Florestal Brasileiro pela concessao das amostras e sua identificaçao e ao INCTBio pelo apoio financeiro.
REFERENCIAS 1. Wheeler, E.A.; Baas, P.; IAWA J. 1998, 19, 241. DOI: http://dx.doi.org/10.1163/22941932-90001528 2. Duarte, V. C. S.; Oliveira, T. L. C.; Coradin, V. T. R.; Camargos, J. A. A.; Gontijo, A. B.; Marques, L. F.; Pastore, T. C. M.; Resumos da 37ª Reuniao Anual da Sociedade Brasileira de Química, Natal, Brasil, 2014. 3. Pastore, T. C. M.; Braga, J. W. B.; Coradin, V. T. R.; Magalhaes, W. L. E.; Okino, E. Y. A.; Camargos, J. A. A.; Muñiz, G. I. B.; Bressan, O. A.; Davrieux, F.; Holzforschung 2011, 65, 127. DOI: http://dx.doi.org/10.1515/hf.2011.010 4. Braga, J. W. B.; Pastore, T. C. M.; Coradin, V. T. R.; Camargos, J. A. A.; Silva, A. R.; IAWA J. 2011, 32, 175. 5. Skoog, D. A.; Holler, F. J.; Nieman, T. A.; Princípios de análise instrumental, 6ª ed., Bookman: Porto Alegre, 2009, cap 18. 6. Pandey, K. K.; Upreti, N. K.; Srinivasan, V. V.; Wood Sci. Technol. 1998, 32, 68. 7. Muyskens, M.; J. Chem. Educ. 2006, 83, 132. DOI: http://dx.doi.org/10.1021/ed083p132 8. Dyer, S. T.; IAWA Bull. 1988, 9, 98. DOI: http://dx.doi.org/10.1163/22941932-90000472 9. Krishna, S.; Chowdhury, K. A.; Indian For. 1935, 61, 74. 10. Miller, R. B.; A guide to more useful woods of the world, International Wood Collectors Society: Madison, 2007. 11. Teixeira, J. G.; Oliveira, B. R. U.; Latorraca, J. V. de F.; Amodei, J. B.; Gurgel, M. de M.; Braz. J. Wood Sci. 2012, 3, 54. 12. Coradin, V. T. R.; Camargos, J. A. A; Marques, L. F.; Silva Jr, E. R.; Madeiras similares ao mogno (Swietenia macrophylla King.): chave ilustrada para identificaçao anatômica em campo, Serviço Florestal Brasileiro, Brasília, 2009. 13. Botelho, B. G.; Reis, N.; Oliveira, L. S.; Sena, M. M.; Food Chem. 2015, 181, 31. DOI: http://dx.doi.org/10.1016/j.foodchem.2015.02.077 PMID: 25794717 14. Gondim, C. S.; Junqueira, R. G.; Souza, S. V. C; Rev. Inst. Adolfo Lutz 2011, 70, 433. 15. Wise, B. W.; Gallagher, N. B.; Bro, R.; Shaver, J. M.; Windig, W.; Koch, R. S; PLS-Toolbox 6.5 for use with MatlabTM (Manual); EigenVector Research Inc., United States of America, 2006. 16. Barker, M; Rayens, W.; J. Chemom. 2003, 17, 166. DOI: http://dx.doi.org/10.1002/cem.785 17. http://www.dca.iag.usp.br/www/material/akemi/fisicaII/apostila_cap_01.pdf, acessada em Janeiro 2015. 18. Valderrama, P.; Braga, J. W. B.; Poppi, R. J.; Quim. Nova 2009, 32, 1278. DOI: http://dx.doi.org/10.1590/S0100-40422009000500034 |
On-line version ISSN 1678-7064 Printed version ISSN 0100-4042
Qu�mica Nova
Publica��es da Sociedade Brasileira de Qu�mica
Caixa Postal: 26037
05513-970 S�o Paulo - SP
Tel/Fax: +55.11.3032.2299/+55.11.3814.3602
Free access