
 |
 |
 |
 |
 |
Revisão
|
|
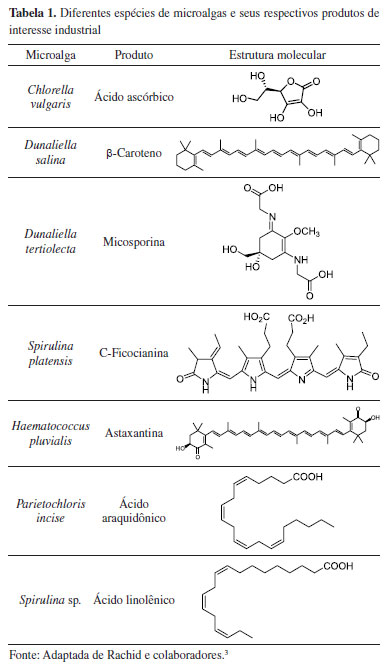
| Microalgas: uma estratégia sustentável na transformação e obtenção de compostos orgânicos Microalgae: a sustainable strategy in the transformation and obtaining of organic compounds |
|
Mauricio Moraes VictorI,II; Fernando Leal Barreiros MoutinhoI,III; Valéria Belli RiattoI,II,*
I. Instituto de Química, Universidade Federal da Bahia, 40170-115 Salvador - BA, Brasil Recebido em 21/06/2023 *e-mail: vriatto@ufba.br The present review presents the biotechnological potential of microalgae, highlighting its applications in biofuels, bioremediation, production of bioactive compounds, biocatalysts, and biofertilizers. Microalgae are photosynthetic microorganisms that combine characteristics of plants and heterotrophic microorganisms, such as yeasts, bacteria, and fungi. They can be considered biotechnological agents of extreme versatility and metabolic adaptability. Its photosynthetic efficiency can absorb large volumes of anthropogenic CO2 and convert it into biomass rich in compounds of high biological and energetic value, enabling the development of sustainable bioremediation processes and the production of third-generation biofuels and biofertilizers. In addition, its biomass has aroused interest in obtaining compounds with biological action, such as carotenoids, mycosporine-like amino acids, polyunsaturated fatty acids, sterols, phycocolloids, carbohydrates, and proteins. The biotechnological potential of microalgae does not end with the production and extraction of its biomass. Its dynamic and adaptive metabolism allows its use in the biotransformation of xenobiotic substrates and the production of compounds of chemical and pharmaceutical interest. In this perspective, microalgae have application qualities as a biotechnological platform in developing synergistic processes between the environment, society, and industry, thus contributing to the expansion and use of Green Chemistry and strengthening the circular economy. INTRODUÇÃO O desenvolvimento de tecnologias capazes de promover a integração de aspectos ambientais, econômicos e sociais tem sido alvo crescente de discussões nas últimas décadas. Esforços coordenados globais, visando o bem-estar do planeta e das futuras gerações, levaram à proposição dos 17 Objetivos de Desenvolvimento Sustentável (ODS) pela Organização das Nações Unidas (ONU).1 Neste contexto, a biotecnologia vem sendo apontada como elemento chave e de grande importância estratégica, devido ao seu potencial diversificado para o uso da biodiversidade na geração de energia renovável, no desenvolvimento de agricultura sustentável e em processos Químicos Verdes.2 Diante deste cenário, as microalgas vêm se destacando em função do seu alto potencial biotecnológico, capazes de produzir diferentes tipos de compostos químicos economicamente importantes, inclusive com relevância comercial.3 A biomassa obtida a partir do cultivo das microalgas é rica em lipídeos e carboidratos, o que permite a produção de biocombustíveis de 3ª geração. Além disso, essa biomassa também é composta por produtos naturais com alto valor agregado, como ácidos graxos poli-insaturados, carotenoides, vitaminas e esteróis. Devido a sua versatilidade metabólica, as microalgas se adaptam a mudanças ambientais e alterações nos meios de cultura, permitindo alta tolerância a fatores abióticos e substratos xenobióticos.4 Estas características, somadas à sua capacidade de cultivo em pequenos espaços e biorreatores, possibilitam controlar seus parâmetros bioquímicos e físico-químicos,5 amplificando seu potencial na obtenção de compostos bioativos. Seu cultivo, em conjunto com a reciclagem de materiais e nutrientes, representa uma oportunidade apreciável, como parte de um sistema econômico regenerativo para exploração de matéria sustentável. As microalgas podem ser consideradas um recurso renovável estratégico, com qualidades para aplicação em diversas áreas biotecnológicas e potencial tendência para aproveitamento em economia circular.6 Embora o emprego das microalgas na produção de biocombustíveis de 3ª geração7 e no tratamento de águas residuais agroindustriais8 esteja bem estabelecido, existem outras aplicações biotecnológicas promissoras que apresentam relevância. A variedade de produtos de alto valor agregado, como fármacos e nutracêuticos, tem despertado interesse do setor industrial (Tabela 1),9 bem como a oportunidade de emprego das microalgas como biocatalisadores de substratos exógenos10 e sua versatilidade como biofertilizantes.11 Em função dessas utilidades destacadas e proeminentes, a presente revisão visa cobrir uma lacuna de divulgação em língua portuguesa, abordando o vasto potencial das microalgas com especial atenção sobre a obtenção de compostos bioativos, desenvolvimento de biofertilizantes e aplicações em biotransformação.
MICROALGAS As microalgas são micro-organismos que abrangem enorme diversidade morfofisiológica e genética (compreendendo seres procarióticos, representados pelas cianobactérias, e seres eucarióticos, representados pelas clorófitas).5 A Figura 1 apresenta uma imagem ilustrativa de microalgas da espécie Dunaliella salina.
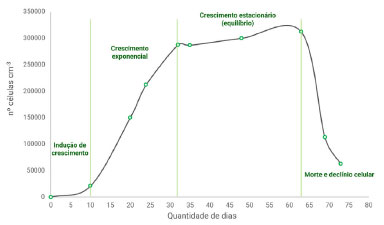 Figura 1. Células de microalgas D. salina obtidas a partir da captura de imagem em equipamento microscópio óptico OPTON modelo TNB-01B com aumento de 400× (fonte: elaborada pelo autor)
O crescimento desses micro-organismos depende principalmente de três fatores: energia, água e fonte de carbono. Seu ciclo de vida pode ser descrito em quatro fases: (i) lag - fase de adaptação fisiológica às condições do meio de cultura e indução de crescimento, com atividade metabólica intensa; (ii) log - crescimento exponencial e multiplicação das microalgas, caracterizada pela presença de células jovens e por ser a fase bioquimicamente mais rica, com a maior atividade metabólica; (iii) estacionária - redução do crescimento, seguida de um pequeno acréscimo até seu valor máximo, caracterizada pelo alto volume de biomassa; (iv) morte celular - fase em que o número de células mortas excede o número de células novas, advém do esgotamento de nutrientes decorrente da alta densidade celular (Figura 2).5
 Figura 2. Curva de crescimento da microalga D. salina em sistema fechado fotoautotrófico, à 25 ºC, com injeção de ar atmosférico e fotoperíodo constante, ao longo de 75 dias (fonte: elaborada pelo autor)Curva de crescimento da microalga D. salina em sistema fechado fotoautotrófico, à 25 ºC, com injeção de ar atmosférico e fotoperíodo constante, ao longo de 75 dias (fonte: elaborada pelo autor)
A principal via metabólica das microalgas é o processo de fotossíntese oxigênica, denominado como mecanismo fotoautotrófico, que utiliza luz como fonte de energia e converte dióxido de carbono em compostos orgânicos, liberando oxigênio molecular a nível celular.12 De forma geral, este processo compreende duas fases: reação fotoquímica (fase clara) e reação bioquímica (fase escura). A fase clara é composta por duas etapas principais, a redução do NADP+ à NADPH no fotossistema I, e a oxidação da água para produção de oxigênio, pelo fotossistema II; sendo estas etapas interligadas por um sistema redox de transporte de elétrons.12 O ATP e o NADPH, produzidos na fase clara, fornecem energia necessária para a fixação do dióxido de carbono no citoplasma, através do ciclo de Calvin, quando o dióxido de carbono absorvido é convertido em carboidrato. Essa síntese acontece pela reação de carboxilação do intermediário ribulose-1,5-bisfosfato, via ação enzimática da ribulose-1,5-bisfosfato carboxilase oxigenase (RuBisCO) (Figura 3).13
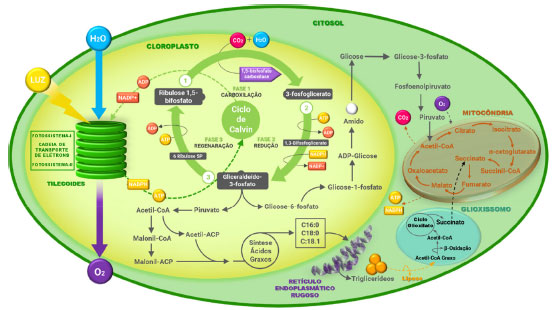 Figura 3. Ilustração do processo metabólico da fotossíntese oxigênica em microalgas (fonte: adaptado de Sen e Yadav, e Bones e colaboradores)12,13
Embora a principal via de crescimento das microalgas seja fotoautotrófica, sua adaptabilidade metabólica possibilita outros sistemas de obtenção de energia e nutrição. No regime heterotrófico, compostos orgânicos são empregados como fonte de energia e fontes de carbono. Já no regime mixotrófico, a energia é obtida através da fotossíntese ou da oxidação de compostos orgânicos.5 A adaptabilidade das microalgas, sob diferentes regimes de cultivo, desponta como uma grande vantagem para aplicação em biotecnologia. A regulação metabólica, imposta nestes meios, permite direcionar e incrementar a obtenção de bioprodutos de interesse (Tabela 2).14

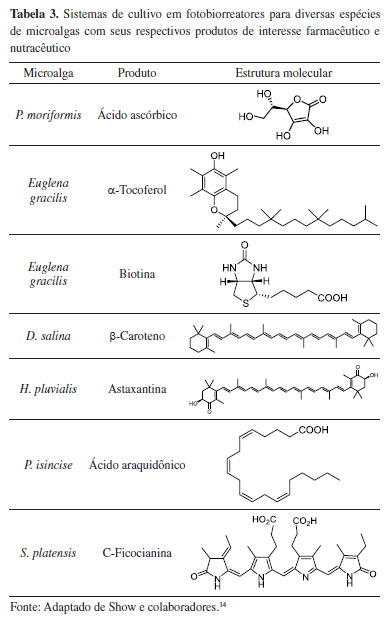
As microalgas também podem ser cultivadas em fotobiorreatores fechados, sendo uma estratégia útil para controlar seus parâmetros bioquímicos e físico-químicos. Estes sistemas podem apresentar configurações diferentes (Figura 4)14 e permitem produzir culturas com elevados graus de pureza, curtos ciclos de cultivo, alta densidade celular, elevadas taxas de biofixação de CO2 e eficiência fotossintética. Os fotobiorreatores são geralmente empregados na produção de bioprodutos de alto valor agregado, tais como farmacêuticos e nutracêuticos (Tabela 3).14
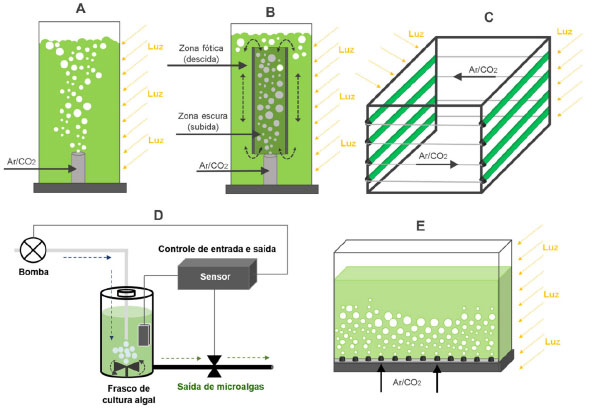 Figura 4. Principais configurações de fotobiorreatores: (A) coluna de bolhas; (B) coluna airlift; (C) tubular horizontal; (D) tanque mecanicamente agitado com bomba de fluxo contínuo; (E) placa plana com tubulação perfurada (fonte: adaptado de Show e colaboradores)14
Dentre as técnicas utilizadas para aprimoramento das culturas de microalgas, a otimização da fixação de carbono nas células, a partir da regulação de suas vias metabólicas, permite a obtenção de bioprodutos, a partir de sua biomassa.15 O uso de compostos químicos como potencializadores na modulação do metabolismo das microalgas, tem demonstrado resultados em ações de regulação de vias biossintéticas, indução de respostas ao estresse oxidativo e de precursores metabólicos (Tabela 4).16 Um exemplo de bioproduto com produção modulada é o ácido docosa-hexaenoico (DHA) (Figura 5), um dos ácidos graxos presentes no ômega-3, que tem atraído grande interesse da indústria de nutracêuticos devido à sua ação neuroprotetiva. Estudos sobre culturas de Chlorella sorokiniana, tratadas com ácido-1-naftalenoacético, associado ao ácido-2-fenilacético, demonstraram aumento no crescimento de sua biomassa rica em lipídios em 104%, enquanto estudos sobre a adição de ácido málico ao meio de cultura de Schizochytrium sp. aumentaram o conteúdo de DHA entre 35-60% em sua composição graxa total (Tabela 4).16
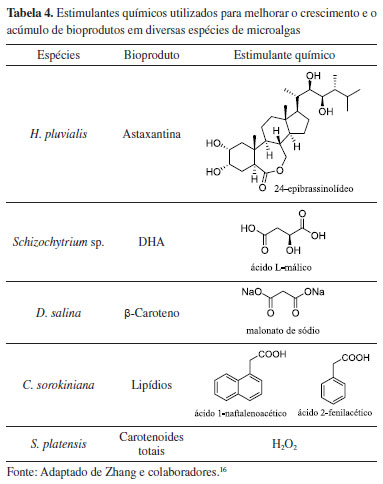
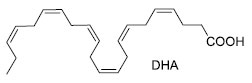 Figura 5. Estrutura do DHA
Assim sendo, o cultivo das microalgas para aplicações biotecnológicas é um processo que depende da fisiologia microbiana e dos sistemas de cultivo. O direcionamento para formação dos bioprodutos pode ocorrer através de duas estratégias: regulação da fisiologia celular, por meio de alterações externas (como pH, fontes de luz, temperatura, adição de nutrientes, etc. ); e engenharia metabólica, através de regulações que compreendem alterações intracelulares das reações fotoquímicas e dos fluxos de carbono e energia das vias metabólicas na mitocôndria e cloroplasto, conforme ilustrado na Figura 6.13 Portanto, o aprimoramento da produção microalgal vem sendo considerado a fim de melhorar e intensificar a composição do conteúdo da sua biomassa e otimizar seu emprego em processos biotecnológicos.
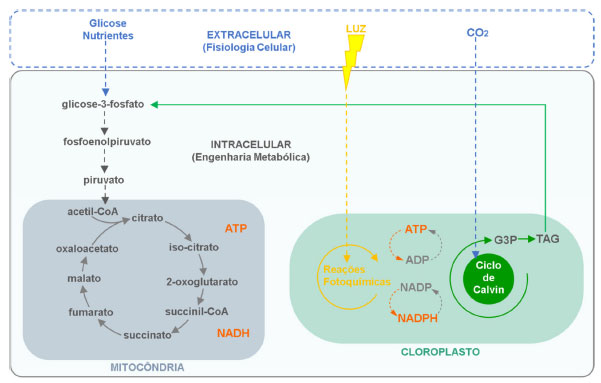 Figura 6. Ilustração das condições extracelulares, ligadas à fisiologia celular, e das condições intracelulares (engenharia metabólica) com os ciclos de reações metabólicas na mitocôndria e cloroplasto (fonte: adaptado de Bones e colaboradores)13
Aplicações biotecnológicas das microalgas Biocombustíveis Biocombustível é um combustível de origem biológica não fóssil, obtido de fontes renováveis e inesgotáveis. Normalmente é produzido a partir de uma ou mais plantas com grande potencial econômico. É uma alternativa viável para substituição do petróleo e combustíveis de origem fóssil não-renovável, com uma série de vantagens ambientais, econômicas e sociais. Contudo, a produção de biocombustíveis a partir de óleos comestíveis apresenta limitações, uma vez que existe competição pela matéria-prima como fonte alimentar. Em função do alto teor lipídico da biomassa microalgal, biorefinarias baseadas em microalgas são utilizadas para geração de bioenergia, sendo a produção do biodiesel um dos principais expoentes.17 Os biocombustíveis provenientes da biomassa de microalgas são classificados como de 3ª geração. O teor de lipídios, carboidratos e proteínas presentes em suas células faz da biomassa microalgal uma matéria prima sustentável e versátil, assim como o aproveitamento de seus coprodutos, para produção e conversão em diferentes fontes de combustíveis renováveis (bioetanol, biogás, bio-óleo e bio-hidrogênio), por meio de diferentes processos químicos, bioquímicos e termoquímicos (Figura 7).18
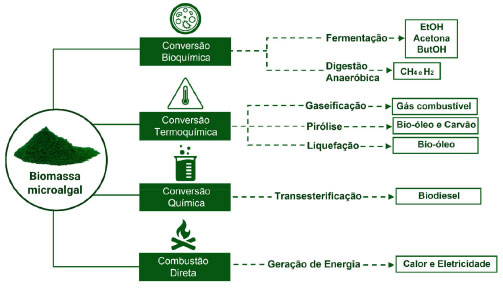 Figura 7. Processos de conversão da biomassa microalgal em diferentes biocombustíveis (fonte: adaptado de Konour)18
A produção de biodiesel de 3ª geração tem se mostrado um recurso energético sustentável e viável, uma vez que, diferentemente das oleaginosas comestíveis (palma, soja, girassol, etc. ), as microalgas não são usadas para alimentação, não necessitam de terras para seu cultivo e possuem sistemas de cultivo adaptáveis e versáteis, com taxas de crescimento rápidas, associadas a uma alta produtividade lipídica.19 Várias espécies de microalgas foram identificadas como produtoras de lipídios para a produção de biodiesel, como Chlorella sp. , C. vulgaris,C. sorokiniana,Chlorella protothecoides,S. platensis,Nannochloropsis gaditana e Nannochloropsis salina.19 De maneira análoga às espécies oleaginosas, o conteúdo lipídico proveniente da biomassa microalgal é constituído pela distribuição graxa de ácidos saturados e insaturados, além da presença de acidos poli-insaturados como o ômega 3 e o ômega 6.5 A produção de biodiesel, a partir de biomassa microalgal, requer um número maior de etapas, quando comparada ao método convencional empregado para oleaginosas (extração de lipídios por prensagem mecânica ou solvente químico, seguida de reação de transesterificação). De forma geral, a produção de biodiesel a partir de microalgas é composta pelas seguintes etapas: colheita da biomassa úmida, secagem, ruptura da parede celular microalgal, extração da biomassa seca por solvente, e produção do biodiesel por meio de reação de transesterificação, na presença de um álcool (metanol ou etanol) e um catalisador (ácido ou basico) (Figura 8).19
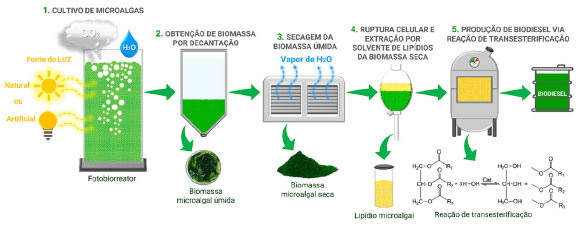 Figura 8. Processo de cultivo e extração da biomassa para produção de biodiesel microalgal (fonte: adaptado de Lam e colaboradores)19
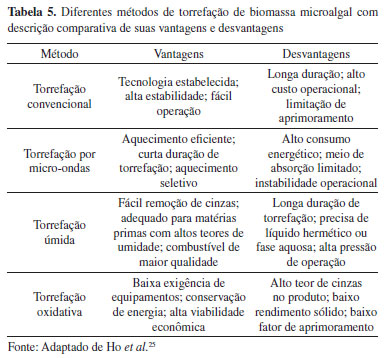
A etapa de extração dos constituintes lipídicos da biomassa microalgal representa um dos principais desafios para as biorefinarias, visto que os rendimentos podem ser baixos e os custos elevados. O processo de ruptura celular (através de ultrassom, agitação mecânica, extração por solvente, etc. ) é a etapa fundamental para a obtenção do conteúdo lipídico de forma satisfatória. Neste contexto, a criação de condições favoráveis de ruptura celular e extração lipídica vem ganhando atenção na literatura.18 Foi realizado um estudo para otimização da extração lipídica da biomassa da microalga Scenedesmus sp. , através da análise de alterações experimentais empregando diferentes misturas de solventes, tempos de extração e formas de ruptura celular.20 A importância destes efeitos foi calculada a partir de uma análise de variância (ANOVA). Segundo os resultados obtidos, o parâmetro que apresentou melhor performance foi a mistura de solventes CHCl3:MeOH (2:1), assistida por ultrassom. Foi observado que duas horas de extração se mostraram suficientes para a obtenção de um rendimento lipídico total de aproximadamente 80 mg g-1 de biomassa seca. Além dos resultados satisfatórios sobre o conteúdo lipídico total obtido, a otimização multivariada apresentou-se como um método acessível, reduzindo o número de experimentos e minimizando o uso de reagentes e energia. A reação de transesterificação in situ (transesterificação direta) tem sido reconhecida como uma alternativa promissora para produção de biodiesel a partir de microalgas.19 Esta metodologia é baseada em um processo one-pot, onde a biomassa (seca ou úmida), o álcool e o catalisador são inseridos no meio reacional. Em uma única etapa, os lipídios são extraídos e convertidos aos respectivos ésteres de ácidos graxos. Idris e colaboradores21 realizaram estudos com a microalga Nannochloropsis sp. , para produção de biodiesel por meio da transesterificação direta (in situ), via catálise ácida (H2SO4), utilizando metanol e líquido iônico (metil sulfato de 1-etil-3-metilimidazol [EMIM][MeSO4]), assistida por aquecimento em micro-ondas. Os resultados demonstraram que a transesterificação direta converteu os lipídios em seus respectivos ésteres metílicos de ácidos graxos sem a necessidade das etapas de secagem e extração, mostrando-se mais efetiva quando comparada ao processo em duas etapas. Os resultados revelaram um rendimento de 41% de biodiesel por biomassa seca, enquanto a composição dos ésteres metílicos graxos apresentou alto teor de insaturações, mostrando ser adequada para a produção de biodiesel. Embora as microalgas apresentem alto conteúdo lipídico em sua biomassa, a aplicabilidade na produção de biodiesel depende da sua composição graxa (comprimento da cadeia, grau de insaturação e presença de ramificações), dado que estes fatores impactam diretamente nas propriedades finais do biodiesel obtido. A avaliação da produção de biodiesel, a partir do biossólido gerado pela ação das microalgas Chlorella sp. ,Euglena sp. e Phacus sp. , em águas de tratamento de efluentes, demonstrou que o biodiesel possuía um teor de 79,5% de ésteres metílicos de ácidos graxos, valor abaixo do mínimo exigido pela Agência Nacional do Petróleo, Gás Natural e Biocombustíveis (ANP) (96,5%).22 De acordo com este estudo, o conteúdo lipídico total do biossólido continha outros compostos (octa-, hexa- e heptadecano), responsáveis por influenciar negativamente a densidade e a viscosidade do biodiesel produzido. Bioetanol e biobutanol também podem ser obtidos através da biomassa microalgal. A produção do bioetanol se assemelha aos processos de conversão de materiais lignocelulósicos, nos quais é empregada hidrólise ácida ou enzimática, permitindo a conversão da biomassa nos açúcares necessários para o processo fermentativo. A limitação de nitrogênio ao meio de cultivo é uma técnica importante que permite o acúmulo de carboidratos e otimização na produção de bioetanol.23 Estudos realizados com Synechococcus sp. registraram um acúmulo de 60% do conteúdo de carboidratos (3 g L-1 de concentração de biomassa), sob condições de restrição de nitrato. Tratamento da biomassa com lisozima, seguido de fermentação com a levedura Saccharomyces cerevisiae, produziu bioetanol em 86% de rendimento. Já a produção de biobutanol é realizada via um processo de fermentação conhecido como acetona-butanol-etanol (ABE). Este processo é constituído por duas etapas. Na primeira, chamada via acidogênese, bactérias do gênero Clostridium realizam a conversão anaeróbica dos carboidratos presentes na biomassa microalgal em ácidos orgânicos voláteis (como ácido acético e ácido butírico). Na segunda etapa, a via solventogênese, estes ácidos são transformados em butanol, acetona e etanol.24 Os biocombustíveis sólidos também vêm se destacando por serem excelentes alternativas aos combustíveis convencionais (fósseis e lenha). Eles possuem alto poder calorífico e emitem menos gases do efeito estufa, podendo ser considerados uma alternativa verde para a redução da dependência energética. A conversão termoquímica da biomassa microalgal é uma estratégia importante para obtenção destes biocombustíveis. O processo de torrefação baseia-se na pirólise branda da biomassa, através de temperaturas entre 200 à 300 ºC, com durações de até 60 minutos. Os sistemas de torrefação podem variar conforme o método de aquecimento e gás de arraste empregados (Tabela 5).25
A torrefação da biomassa microalgal está relacionada diretamente com sua composição. Estudos sobre a decomposição térmica de microalgas demonstraram que as biomassas com alto teor de carboidratos são mais facilmente degradadas, seguidas de biomassas ricas em proteínas e lipídios.26 Desta forma, biomassas com altos teores de carboidratos resultam em maior perda de massa e, consequentemente, baixos valores caloríficos. Entretanto, o aumento no conteúdo proteico da sua biomassa pode conferir incremento significativo no valor calorífico, potencializando assim sua viabilidade como biocombustível sólido. Contudo, a queima do conteúdo proteico pode emitir gases com altos teores de nitrogênio e enxofre, tornando-se necessário manter um equilíbrio entre o potencial energético e a emissão de gases durante o processo de torrefação. Para este efeito, é necessário controlar o teor de proteínas nas células durante seu cultivo. Devido ao desenvolvimento rápido do sequenciamento genético das microalgas, algumas abordagens de edição gênica se desenvolveram consideravelmente nos últimos anos.25 As microalgas e seus componentes celulares podem ser manipulados com precisão, direcionando a formação de compostos orgânicos necessários para otimização da produção de biocombustíveis sólidos, a partir da torrefação (Figura 9). A superexpressão de algumas enzimas-chave para regulação metabólica pode maximizar a produção de amido, proteínas ou inibir vias metabólicas competitivas. Como consequência, o biosólido produzido, após torrefação, possui propriedades superiores de combustível e eficiência energética. Propriedades como valor calorífico, hidrofobicidade, capacidade de moagem, armazenamento e a homogeneidade do combustível podem ser aprimoradas significativamente.25
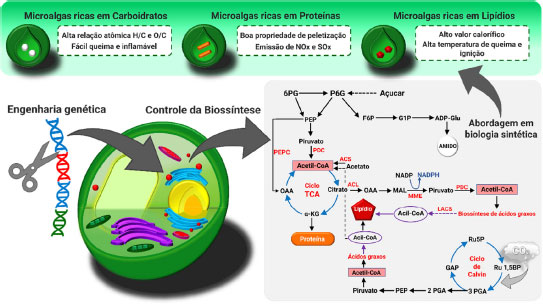 Figura 9. Edição de genes em microalgas para aprimorar a obtenção de biocombustíveis sólidos (fonte: adaptado de Ho et al.)25
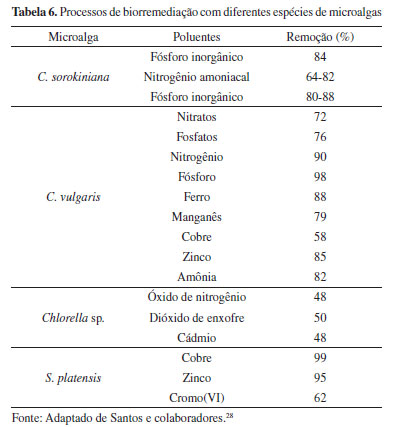
Biorremediação O potencial de adaptação das microalgas às mudanças das condições ambientais, através de seus mecanismos de aclimatação bioquímica e fisiológica, possibilita seu crescimento em ambientes aquáticos eutrofizados. Seu metabolismo viabiliza processos de biossorção de íons metálicos, qualificando-as como uma alternativa biotecnológica eficiente e sustentável para emprego em técnicas e projetos de biorremediação (Tabela 6).27,28
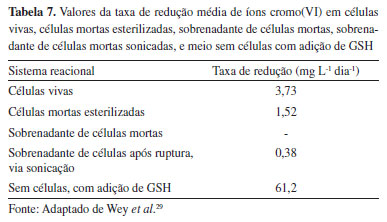
O cromo é um metal pesado frequentemente encontrado no meio ambiente. Entre suas formas iônicas predominantes, o cromo(III) tem apenas cerca de um milésimo da toxicidade do cromo(VI). Em geral, a biomassa de microalgas pode ser empregada como adsorvente durante o processo de remoção de cromo. Contudo, estudos sobre a aplicação da microalga C. vulgaris na biorremediação de íons cromo(VI) mostraram a conversão do cromo(VI) em cromo(III) através de dois mecanismos sinérgicos: (i) mecanismo de redução biológica, via enzima cromo redutase; e (ii) mecanismo via oxidação da glutationa (GSH), proveniente da ruptura celular.29 Este fenômeno foi observado quando culturas de células microalgais mortas e esterilizadas foram capazes de reduzir o cromo(VI). Foram realizados ensaios para avaliar o potencial de redução do cromo(VI) empregando: (i) células microalgais vivas; (ii) células mortas esterilizadas; (iii) sobrenadante do meio de células mortas; (iv) sobrenadante do meio de células após ruptura por sonicação; e (v) meio sem células, com adição de GSH e cromo(VI). Os resultados demonstraram a influência da glutationa presente no meio celular para redução do cromo(VI) em cromo(III) (Tabela 7).
Associado a processos de biorremediação, a aplicação de microalgas na biodegradação de contaminantes emergentes como fármacos, hormônios, retardantes de chama e herbicidas, vem se mostrando uma importante opção biotecnológica para tratamento de efluentes e águas costeiras.17 Estudos sobre poluentes endócrinos presentes em sistemas marinhos costeiros demonstraram a habilidade das microalgas D. salina, Nannochloropsis oculata, Platymonas subcordiformis e Phaeocystis globosa de degradar o nonilfenol etoxilado, um importante contaminante ambiental que oferece riscos à saúde e ao meio ambiente.30 Em outro estudo, foi avaliado o potencial de biorremediação da microalga Selenastrum capricornutum em efluentes e águas residuais, para contaminação com hormônios esteroidais. Os resultados apresentaram um potencial de remoção e biotransformação dos hormônios 17β-estradiol e 17α-etinilestradiol em 93,9 e 75,3%, respectivamente.31 Estudos também demonstraram a versatilidade das microalgas em interagirem com outros micro-organismos, promovendo processos biotecnológicos.32 Um sistema de célula combustível microbiano (CCM), utilizando S. cerevisiae e S. platensis, foi empregado na geração simultânea de energia renovável e produção de biomassa, a partir de águas residuais oriundas de indústrias cafeeiras. Sistemas bioeletroquímicos de CCM são dispositivos capazes de converter matéria orgânica em energia elétrica pela ação de micro-organismos. A matéria orgânica contida em águas residuais foi utilizada pelos micro-organismos como nutriente de crescimento, gerando como consequência processos metabólicos capazes de fornecer prótons e elétrons que foram convertidos em energia elétrica (Figura 10). A redução do conteúdo orgânico de águas residuais industriais ocorreu no ânodo da CCM, pela ação enzimática da levedura S. cerevisiae, tendo como produto principal o CO2. No cátodo, a microalga S. platensis consumiu o CO2, via fotossíntese, regenerando o O2. Este sistema gerou um fluxo de elétrons através do sistema redox bioeletroquímico da CCM, produzindo uma corrente elétrica de 0,17 V, redução de 60% da demanda química de oxigênio, além da redução de 83% dos resíduos sólidos totais.
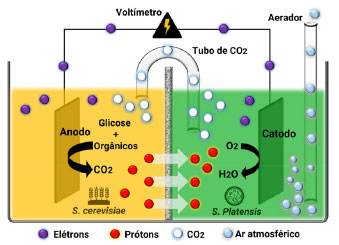 Figura 10. Esquema da célula combustível integrada utilizando a levedura S. cerevisiae e a microalga S. platensis (fonte: adaptado de Christwardana et al.)32
Produção de compostos bioativos As microalgas são candidatas promissoras a plataformas de produção de compostos orgânicos de alto valor agregado. Elas apresentam várias vantagens intrínsecas: são seres eucarióticos, capazes de realizar funções biológicas e bioquímicas complexas; são fotossintetizantes, portanto têm requisitos de crescimento baratos; são resistentes a estresses associados ao cultivo industrial; e podem ser apropriadas para produção em escala industrial. As tendências atuais de biotecnologia envolvendo microalgas propõem a obtenção de compostos bioativos de alto valor agregado, integrados a sistemas produtivos de biocombustíveis e processos de biorremediação. Estes processos envolvem o emprego da biomassa microalgal em grande escala, de forma sustentável e economicamente viável.33 Dentre os compostos orgânicos provenientes das microalgas, os mais explorados são triacilglicerídeos (TAGs), carotenoides e ácidos graxos poli-insaturados de cadeia longa (LC-PUFAs). Dentre esses últimos, destacam-se os ácidos graxos ômega-3 eicosapentaenóico (EPA) e docosa-hexaenóico (DHA), empregados em suplementação nutricional humana e que atraem grande atenção da indústria de nutracêuticos.33 A obtenção destes importantes ácidos graxos através das microalgas é bastante promissora, podendo tornar-se uma alternativa economicamente mais interessante em detrimento de suas fontes habituais, como peixes, farinhas de sementes e castanhas ou óleos. Nesta perspectiva, estudos sobre produção de EPA em águas residuais industrias de celulose e papel, em efluente de digestão anaeróbica, a partir do cultivo da microalga N. oculata, apresentaram resultados interessantes de rendimento (47,2 mg de EPA por litro de cultura), além do aumento de aproximadamente sete vezes mais produção de TAGs, quando comparados a regimes de cultivo em água do mar artificial.34 Avanços recentes em tecnologias de sequenciamento, metabolômica e proteômica têm pavimentado o caminho para a caracterização metabólica a nível genético de algumas espécies de microalgas. A possibilidade de direcionar geneticamente a produção de compostos orgânicos, colocou essas espécies sob os holofotes como biofábricas promissoras de tamanho celular, movidas à energia solar.35 A biossíntese e o acúmulo de compostos específicos dependem, no entanto, da genética da espécie da microalga, fase de crescimento, disponibilidade de nutrientes, salinidade, temperatura e irradiação. Estratégias para melhorar a produção de triacilglicerídeos, ácidos graxos poliinsaturados de cadeia longa e carotenoides tem sido empregadas em cepas selecionadas e aprimoradas por via de engenharia metabólica (por exemplo, mutagênese direcionada). Esses processos podem resultar em microalgas que exibem taxas de crescimento mais aceleradas, robustez contra mudanças ambientais e contaminantes, bem como a capacidade de produzir carotenoides, TAGs e LC-PUFAs simultaneamente (Figura 11).33
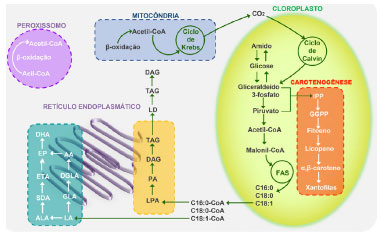 Figura 11. Esquema simplificado dos compartimentos e das vias metabólicas da carotenogênese, síntese de ácidos graxos e de triacilgliceróis em microalgas (fonte: adaptado de Varela e colaboradores)33
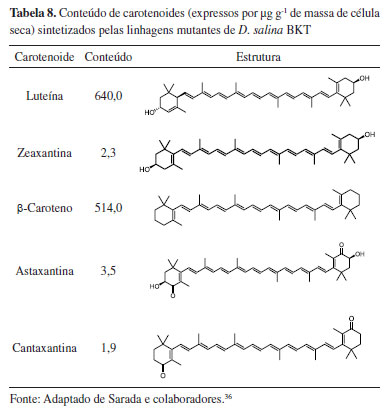
A biossíntese de TAGs e LC-PUFAs ocorre na membrana do retículo endoplasmático. Alguns exemplos de LC PUFAs são os ácidos linoleico (LA), α-linolênico (ALA), γ-linolênico (GLA), estearidônico (SDA), araquidônico (AA), di-homo-γ-linolênico (DGLA), eicosatetraenóico (ETA), EPA e DHA. Também no retículo endoplasmático o glicerol-3-fosfato é esterificado na presença de três ácidos graxos, gerando TAGs, incluindo os intermediários lisofosfatidato (LPA), ácido fosfatídico (PA) e diacilglicerol (DAG). Embora a topologia da biossíntese de carotenoides não seja totalmente conhecida, as evidências atuais sugerem que esta ocorra principalmente no cloroplasto. A carotenogênese começa com o pirofosfato de isoprenila (IPP), que é sintetizado a partir de piruvato e gliceraldeído-3-fosfato. Após várias etapas de condensação, o pirofosfato de geranilgeranila (GGPP) é convertido em fitoeno, o primeiro carotenoide a ser sintetizado. As reações de dessaturação levam ao licopeno, que pode seguir duas rotas diferentes, produzindo α- ou β-caroteno. Sua hidroxilação pode dar origem a compostos de alto valor agregado como as xantófilas (por exemplo, luteína e astaxantina). Explorando essa alternativa tecnológica, a microalga D. salina foi estudada como modelo para produção de cetocarotenoides a partir de engenharia metabólica da via biossintética de carotenoides.36 A expressão da enzima β-caroteno-cetolase (BKT) foi realizada através da introdução do gene bkt da microalga H. pluvialis. Os resultados obtidos demonstraram a viabilidade na produção de luteína e outros carotenoides a partir das linhagens recombinantes de D. salina (Tabela 8).
Terpenos são compostos naturalmente produzidos pelas plantas, através de seu metabolismo secundário. São frequentemente biossintetizados em pequenas quantidades ou em misturas de compostos semelhantes, o que torna a extração das fontes naturais comercialmente inviável. A síntese química também é desafiadora, devido à complexidade das estruturas de interesse. Geralmente são comercializados na forma de óleos essenciais e estão relacionados a uma grande variedade de propriedades biológicas, como anticancerígena, anti-inflamatória, bactericida e fungicida. A biossíntese de terpenoides possui duas vias metabólicas, a via do metileritritol-fosfato (MEP), que ocorre no cloroplasto; e a via do mevalonato (MVA), situada no citosol, no peroxissoma e no retículo endoplasmático. Ambas vias metabólicas possuem dois precursores, o pirofosfato de isoprenila (IPP) e o pirofosfato de dimetilalila (DMAPP), que atuam como unidades elementares na biossíntese dos isoprenoides (Esquema 1).37 A via MVA ocorre em Archaea e no citosol da maioria dos organismos eucarióticos. Começa com acetil-CoA e termina com a formação de IPP, que é interconvertido em seu isômero alílico DMAPP pela ação da IPP isomerase (IDI). A via MEP está presente na maioria das bactérias, em plastídeos de organismos fotossintéticos (como as microalgas) e em alguns outros organismos eucarióticos. Ao contrário da via MVA, a via MEP é alimentada por piruvato e gliceraldeído-3-fosfato (GAP). Seus produtos finais, IPP e DMAPP, são simultaneamente sintetizados a partir de seu precursor comum, 4-hidróxi-3-metilbut-2-enil-difosfato (HMBPP), e a enzima IDI isomerase converte o excesso de IPP em DMAPP. Então, IPP e DMAPP são condensados para formar o precursor dos monoterpenos, pirofosfato de geranila (GPP). Consecutiva adição de unidades IPP forma o pirofosfato de farnesila (FPP) e o pirofosfato de geranilgeranila (GGPP), precursores de sesquiterpenos e diterpenos, respectivamente. Pirofosfatos de prenila com cadeias de carbono mais longas (C30 e C40) também são produzidos. A última parte da biossíntese, que fornece uma vasta diversidade de terpenoides, ocorre após transformações e modificações dos esqueletos mencionados através de enzimas, como oxido-redutases, hidrolases, transferases, etc.
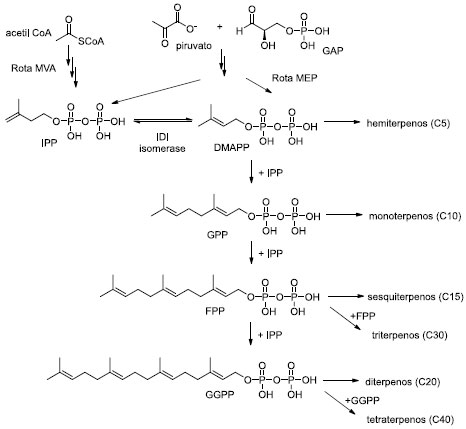 Esquema 1. Biossíntese de isoprenoides via rota do mevalonato (MEV) e na via rota metileritritol-fosfato (MEP) (fonte: adaptado de Vavitsas et al.)38
Associando a produção promissora de terpenos pelas microalgas, com a possibilidade de potencializá-la através de ferramentas de engenharia genética, estudos envolvendo expressão gênica em algumas espécies vêm se destacando.38 A microalga Chlamydomonas reinhardtii é um organismo modelo amplamente estudado em genética de algas. Os esforços da engenharia desta espécie mostraram sua adequação como uma fábrica unicelular para a síntese de pequenas moléculas provenientes de enzimas fruto de edição. A microalga foi manipulada para produzir o sesquiterpeno (E)-α-bisaboleno (Figura 12).39 Por meio da engenharia genética, melhorias substanciais da sua produtividade foram alcançadas através do ajuste coordenado do metabolismo de isoprenoides, combinando com a superexpressão da enzima terpeno sintase. Até 10,3 mg de bisaboleno por grama de peso seco de célula foram produzidos em cinco dias, o que representou um aumento de mais de quinze vezes em relação às cepas de expressão padrão de sintase. A investigação do desempenho da cepa em cultivos de aumento de escala determinou o benefício de produtividade do bisaboleno nos ciclos claro:escuro. O cultivo mixotrófico rendeu a formação de bisaboleno em até 11 mg L-1 de cultura, em sete dias. Através do regime fotoautotrófico, o bisaboleno foi obtido em apenas 3,9 mg L-1 de cultura.
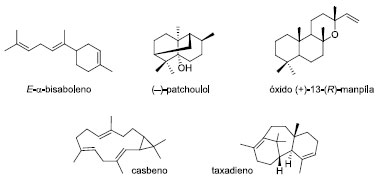 Figura 12. Isoprenos produzidos através de linhagens de microalgas C. reinhardtii recombinantes
A microalga C. reinhardtii também apresentou potencial para produzir o sesquiterpenoide patchoulol (Figura 12).40 Através da expressão da enzima patchoulol sintase, foi avaliada a produção do sesquiterpeno em condições diferentes de crescimento da microalga. Em condições fotoautotróficas, foi observada a produção do patchoulol em até 1,03 mg L-1 de cultura. Em outro estudo, a expressão de enzimas vegetais heterólogas do genoma nuclear da microalga C. reinhardtii potencializou a produção dos diterpenoides casbeno, taxadieno e óxido de (+)-13-(R)-manoíla (Figura 12).41 Através de cultivos fotoautotróficos, usando CO2 como única fonte de carbono, ciclos de iluminação dia:noite e sete dias de cultivo, foram produzidos até 80 mg de óxido de (+)-13-(R)-manoíla por grama de massa seca de células. Essas conquistas representaram um marco importante na engenharia da microalga C. reinhardtii em direção à perspectiva de projetar biofábricas sustentáveis de microalgas movidas à luz. Embora apenas estudos restritos envolvendo algumas espécies de microalgas vêm sendo relatados, estes micro-organismos possuem diferentes características morfológicas, fisiológicas e genéticas, o que lhes confere grande capacidade de produzir diferentes metabólitos biologicamente ativos. A biotecnologia das microalgas tem se tornado objeto de estudo em diversas áreas devido à variedade de bioprodutos que podem ser obtidos. Quando seus processos de cultivo são melhor compreendidos, as microalgas podem se tornar uma fonte ambientalmente correta e economicamente viável de produção de compostos de alto valor agregado. Biocatálise A redução de compostos carbonílicos tem sido explorada extensivamente através de biotransformações.42 O emprego de micro-organismos heterotróficos em reações de biorredução assimétrica está bem estabelecido,43 contudo a utilização de organismos fotoautotróficos, como as microalgas, é relativamente limitada.44 O método de cultivo das microalgas é distinto daquele usado para micro-organismos heterotróficos e, portanto, são raros os químicos orgânicos síntéticos que estão familiarizados com os organismos fotoautotróficos. Nas reações catalisadas por enzimas redutases, o hidreto transferido é fornecido pelos cofatores nicotinamida adenina dinucleotídeo (NADH) ou seu respectivo fosfato (NADPH). Esses cofatores são proibitivamente caros se usados em quantidades estequiométricas e sua regeneração in situ, através de uma segunda reação redox, permite que sejam inseridos novamente no ciclo da reação. As microalgas combinam características metabólicas microbianas (crescimento rápido e cultura de alta densidade) com as qualidades das plantas (fotossíntese autotrófica). Estas propriedades possibilitam seu emprego como fonte de células biocatalíticas na síntese enzimática e assimétrica de produtos quirais.45 Seu metabolismo fotossintético possui fotoenzimas, enzimas que requerem um fluxo constante de luz para catalisar uma reação química. Durante a fotossíntese, ocorre a regeneração do cofator NADP+ em NADPH através do fluxo de elétrons em seus fotossistemas I (PSI) e II (PSII) (Figura 13).46 Estes fotossistemas permitem a conversão da energia luminosa em energia química (NADPH e ATP) e a produção de oxigênio molecular. A primeira etapa envolve a foto-oxidação da água em oxigênio molecular, a geração de dois elétrons e a liberação de dois prótons pelo PSII. Então, o PSII canaliza os elétrons liberados a uma plastoquinona (PQ). Este processo fornece o gradiente de prótons necessário para a síntese de ATP. Os elétrons (e prótons) são canalizados através de uma cadeia de transferência de elétrons ao complexo de citocromo b6f (Cytb6f), que libera os prótons para o lúmen (auxiliando na geração do gradiente de prótons) e transfere os elétrons para o PSI. No PSI ocorre a transferência de elétrons através da ferredoxina, regenerando o NADPH pela enzima ferredoxina-NADP+ redutase (FNR).
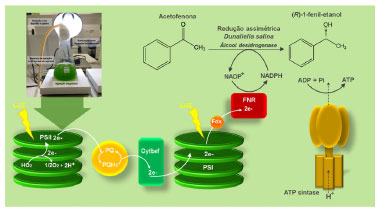 Figura 13. Metabolismo fotossintético na regeneração de cofatores NADP+ em NADPH, através do fluxo de elétrons em seus fotossistemas I e II (fonte: adaptado de Kroutil e colaboradores)46
É admissível que o poder redutor do NADPH, gerado através da fotossíntese, também possa ser usado na redução de substratos exógenos, como cetonas pró-quirais não naturais, produzindo álcoois opticamente ativos. Portanto, a autossuficiência enzimática na regeneração de cofatores, associada à sua ação biocatalítica, atribuem às microalgas características fundamentais em biocatálise assimétrica, como altas estereosseletividades e quimiosseletividades, somadas ainda ao fato de serem micro-organismos seguros, ou seja, não são tóxicos e nem patogênicos.44 Estudos sobre o potencial biocatalítico de microalgas com células íntegras de S. platensis e D. salina, tendo acetofenona como substrato modelo, apresentaram, respectivamente, resultados expressivos com 53 e 100% de conversão e 99 e 97% de excesso enantiomérico (ee) para os enantiômeros (S)-1-fenil-etanol e (R)-1-fenil-etanol.47 A diversidade enantiomérica observada para o mesmo substrato, associada às purezas ópticas, demonstrou o potencial destas microalgas como células íntegras em reações de biocatálise assimétrica (Figura 14).
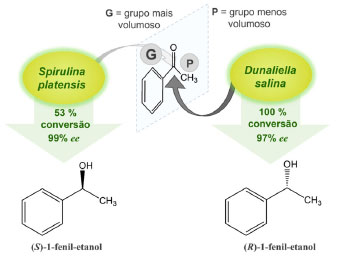 Figura 14. Reações de biocatálise assimétrica com células íntegras de microalgas S. platensis e D. salina
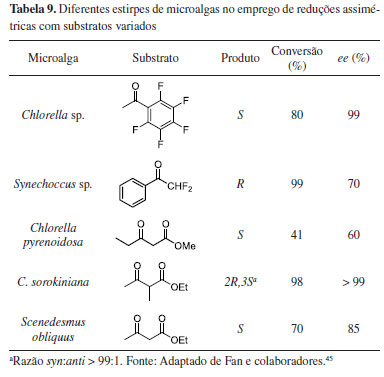
Nesta perspectiva, além da estirpe D. salina, uma variedade de outras espécies de microalgas vem sendo aplicada em estudos de biocatálise assimétrica como fonte de células biocatalíticas. A Tabela 9 ilustra alguns estudos sobre redução assimétrica de cetonas pró-quirais e β-ceto-ésteres.45
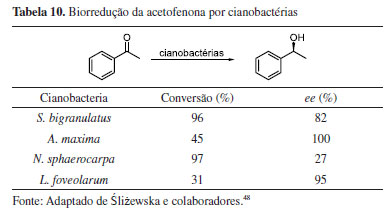
A capacidade biorredutora de quatro cepas de cianobactérias foi avaliada para a redução da acetofenona ao álcool feniletílico quiral correspondente (Tabela 10).48 As condições foram otimizadas com relação à concentração do substrato, duração do período de pré-cultivo, tempo de reação, regime de luz e adição de glicose ao meio de cultura. Os resultados demonstraram que a eficiência da reação aumentou com o tempo de biotransformação. No entanto, o aumento da concentração do substrato limitou o rendimento do processo. Além disso, todas as cepas testadas realizaram a reação com maior eficácia sob regime de luz contínua. As cepas mais ativas, Nodularia sphaerocarpa e Synechococcus bigranulatus, realizaram a conversão de 1 mmol L-1 de acetofenona com conversões de 97 e 96% após 13 dias de biotransformação, respectivamente. O álcool com configuração S foi obtido preferencialmente em todas as reações, com excessos enantioméricos acima de 95% para cepas de Leptolyngbya foveolarum e Arthrospira maxima.
Estudos utilizando as cianobactérias Synechocystis sp. , N. Sphaerocarpa e Synechococcus sp. mostraram suas efetividades como biocatalisadores para redução de carbonilas de compostos exógenos (Tabela 11). A cianobactéria Synechococcus sp. também foi capaz de realizar a redução quimio- e enantiosseletiva da 2-metil-2-ciclopenten-1-ona, gerando a (S)-2-metilciclopentanona em 99% de conversão e 98% de excesso enantiomérico (Tabela 11).49
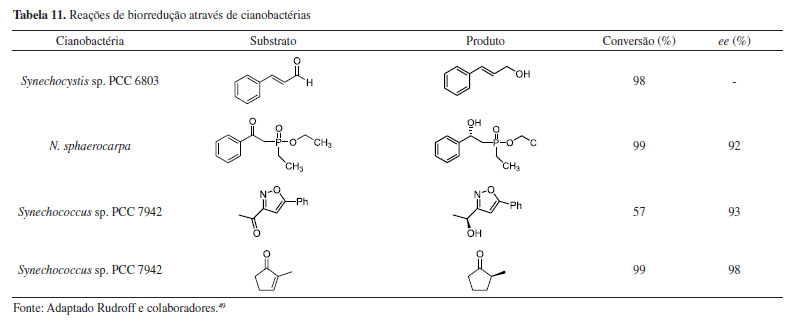
Yang e colaboradores50 realizaram estudos sobre métodos de regulação metabólica na promoção eficiente da redução assimétrica da acetofenona com microalgas da espécie S. platensis. Foram empregados inibidores químicos, como ácido iodoacético e trioxodinitrato de sódio (sal de Angelis), para diminuição da gliceraldeído-3-fosfato desidrogenase (GAPDH) e consequente aumento do NADPH intracelular microalgal. Segundo os autores, este processo ocorre porque a diminuição da GAPDH faz com que reduza o consumo de NADPH no ciclo de Calvin e, consequentemente, aumente o potencial de regeneração de NADPH via fotossíntese, assim como sua capacidade de redução biocatalítica. Em todas as reações, o enantiômero (S)-1-feniletanol foi obtido com excessos enantioméricos maiores que 99%. Os resultados obtidos demonstraram que a adição de 0,10 μmol L-1 de sal de Angelis promoveu incremento do rendimento de 29% para 48%, mas também foi observado que esse aumento é limitado a uma concentração específica para cada agente inibidor, conforme apresentado na Tabela 12.
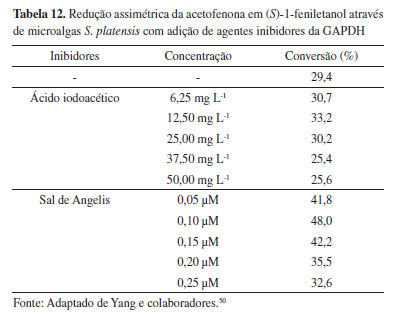
Kojima e colaboradores51 avaliaram a influência do volume do grupo alcóxi na redução assimétrica de β-ceto-ésteres, empregando a cianobactéria Synechocystis sp. PCC 6803, sob iluminação de LED vermelha (660 nm) (Tabela 13). Foi observada a obtenção dos (R)-β-hidróxi-ésteres correspondentes em excessos enantioméricos medianos. O aumento da R-seletividade ocorreu em função do aumento do volume do grupo alcóxi-éster. Posteriormente, o 3-oxobutanoato de terc-butila foi submetido a um segundo estudo para avaliar o efeito da quantidade de células de cianobactérias sobre as reações. A quantidade de células foi ajustada com base na absorbância à 680 nm no espectro de absorção ultravioleta visível, ajustando a absorbância em 750 nm para zero (Abs680-750). Foi observado que as conversões e os excessos enantioméricos aumentaram com a diminuição da concentração do substrato no meio de cultura, sugerindo uma possível toxicidade provocada pelo substrato exógeno para o micro-organismo. Contudo, para uma quantidade de células acima de Abs680-750 0,990, os valores de excesso enantiomérico diminuem, podendo ser uma consequência da desigualdade da exposição das células à luz, chamado de fenômeno da auto-sombra, que ocorre para culturas com alta densidade celular.
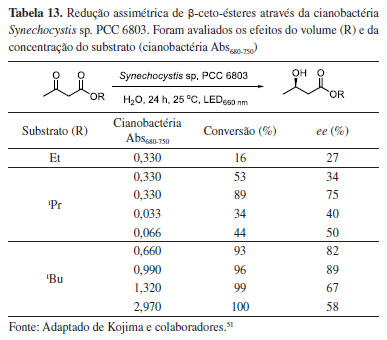
Após o êxito do estudo anterior, Kojima e colaboradores52 avaliaram a regulação metabólica da cianobactéria Synechocystis sp. PCC 6803, mediada pela adição de solventes orgânicos. O estudo foi realizado através da redução assimétrica do 3-oxobutanoato de terc-butila, após 24 h e iluminação de LED vermelha (660 nm). Na ausência de solvente orgânico, o (R)-β-hidróxi-éster correspondente foi obtido em 81% de rendimento e 79% de excesso enantiomérico, enquanto que a adição de tolueno (1% v/v) ao sistema levou à formação do outro enantiômero, o (S)-β-hidróxi-éster correspondente, em 87% de rendimento e excelente estereosseletividade (Esquema 2). Uma S-seletividade também foi observada na presença de clorofórmio, benzeno, etilbenzeno, ciclo-hexano e metilciclo-hexano. Os autores discutem que, embora o mecanismo de adição destes solventes no sistema não esteja muito claro, provavelmente a adição de tolueno interrompe o sistema fotossintético, reduzindo a capacidade regenerativa do cofator NADPH e impedindo a função da redutase R-seletiva, resultando na ativação relativa da redutase S-seletiva. A interrupção do sistema fotossintético deve ser causada pela extração extracelular da clorofila por solventes que exibem maior afinidade por ela. Solventes orgânicos polares, como DMSO, THF e etanol não apresentam tal efeito.
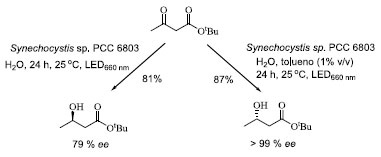 Esquema 2. Efeito da adição de solventes orgânicos na reação de biorredução do 3-oxobutanoato de terc-butila, mediada pela cianobactéria Synechocystis sp. PCC 6803
Devido ao conhecimento adquirido nos últimos anos envolvendo engenharia genética microalgal, a aplicação desta ferramenta também vem sendo empregada para potencializar reações de biocatálise de substratos exógenos. Estudos envolvendo a técnica de expressão enzimática integrada à biotecnologia microalgal demonstraram ser possível a biossíntese de aminas alifáticas em regime one-pot, com reciclagem in situ do cofator NADPH, a partir da enzima amina desidrogenase (EsLeuDH-DM) expressa pela microalga C. reinhardtii (Figura 15).53
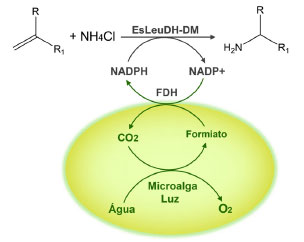 Figura 15. Amina desidrogenase (EsLeuDH-DM) associada à microalga C. reinhardtii, com reciclagem in situ do cofator NADPH (fonte: adaptado de Gröger e colaboradores)53
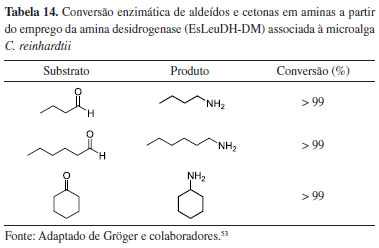
A microalga C. reinhardtii recombinante foi capaz de transformar aldeídos e cetonas alifáticos nas respectivas aminas, em excelentes taxas de conversão (Tabela 14). As reações de biotransformação foram realizadas por meio da enzima amina desidrogenase EsLeuDH-DM, através do processo fotoautotrófico da cultura de C. reinhardtii, combinada com a ação enzimática da formiato desidrogenase (FDH), proveniente da levedura Candida boidinii (cb-FDH). O formiato produzido, via metabolismo microalgal, foi convertido em CO2 pela enzima FDH, via catálise oxidativa, fornecendo os elétrons para regeneração do cofator NAD+ em NADPH, utilizado pela enzima EsLeuDH-DM na bioconversão dos substratos.
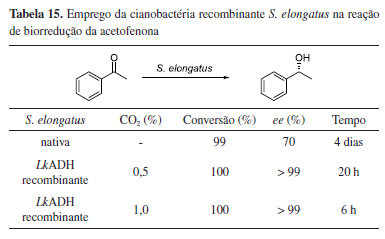
Em um estudo envolvendo engenharia genética de cianobactérias através de uma linhagem de Synechococcus elongatus PCC 7942 recombinante, foi realizada a expressão da enzima álcool desidrogenase LkADH para potencializar a biorredução da acetofenona (Tabela 15).54 Enquanto a reação envolvendo células nativas de S. elongatus forneceu o (R)-1-feniletanol em apenas 70% de excesso enantiomérico em quatro dias de reação,55 o emprego da cianobacteria recombinante, na presença de CO2, elevou o excesso enantiomérico e reduziu o tempo de reação (Tabela 15). Foi observado que o nível de dióxido de carbono desempenha um papel crítico na melhoria da eficiência da bioconversão, possivelmente devido ao aumento do crescimento microalgal e melhor disponibilidade de NADPH. A redução completa da acetofenona em (R)-1-feniletanol, com excesso enantiomérico de 99%, foi alcançada após 6 h de reação e densidade celular relativamente baixa (0,66 g L-1), acoplando luz, níveis de CO2 e sem a necessidade da adição de um co-substrato.
Os resultados aqui relatados expressam os atributos das microalgas como fonte de biocatalisadores. Seu eficiente aparato de fotossíntese, alta taxa de crescimento e acessibilidade à engenharia genética as tornam candidatas promissoras para conduzir reações químicas em escala industrial, em um processo altamente sustentável, capturando gases do efeito estufa e não competindo por terras aráveis. Seu metabolismo fotoautotrófico apresenta características favoráveis para emprego em biocatálise, produzindo produtos quirais com altas enantiosseletividades. Apesar de todas essas vantagens, algumas questões podem limitar as aplicações destes micro-organismos em escala industrial. O aumento de escala de células fotossintéticas requer redesenho dos biorreatores clássicos, baixas densidades celulares e rendimentos insuficientes são observados frequentemente nos grandes biorreatores. Como perspectiva futura, uma maior compreensão da genética destas células admitirá elaborar modelos de engenharia metabólica que permitirão explorar melhor seu potencial. Esta ferramenta possibilitará reduzir reações colaterais indesejadas e redirecionar os recursos para as reações alvo (melhoria da conversão substrato/produto, disponibilidade de cofator otimizada, incremento das estereosseletividades). Finalmente, tendo em vista o grande potencial das microalgas e a crescente necessidade de opções sustentáveis, esforços direcionados para estas questões podem transformá-las numa plataforma real e sustentável para biocatálise assimétrica. Biofertilizantes Os biofertilizantes podem ser empregados como alternativa verde e sustentável aos fertilizantes químicos, restabelecendo a fertilidade típica dos solos e melhorando sua estrutura e função.56 Eles são obtidos a partir de preparações de células microbianas vivas, aumentando o nível nutricional dos solos.57 Seu emprego incrementa o crescimento e a produtividade das plantas por meio de vários mecanismos, como fixação de nitrogênio atmosférico, solubilização e mobilização de fósforo, compostagem, produção de fitormônios e supressão de doenças. Os primeiros quatro mecanismos estão envolvidos no fornecimento de nutrientes para as plantas. Os fitormônios têm um papel crucial no desenvolvimento e diferenciação de tecidos vegetais, e também aumentam a absorção e translocação de micronutrientes (especialmente Fe). Os micro-organismos promotores do crescimento das plantas podem suprimir os patógenos através da produção de antibióticos, enzimas hidrolíticas, sideróforos, HCN e produção de amônia. Estes metabólitos induzem o sistema de defesa da planta e a competição por nutrientes e habitat. A aplicação de biofertilizantes pode aumentar o rendimento da cultura em 20% a 35%, em relação ao controle, e também pode reduzir a aplicação de fertilizantes NPK em até 60%. Estudos sobre as formulações de biofertilizantes baseadas em microalgas têm alcançado destaque no emprego de sua biomassa como fertilizantes verdes alternativos para diversas culturas vegetais, como arroz, tomate, espinafre, milho, batata e cebola (Tabela 16).58 Os resultados observados em diferentes estudos apresentam como principais características: (i) melhoria do rendimento germinativo (altura e comprimento da raiz e da parte aérea); (ii) aumento da taxa de crescimento vegetativo; (iii) aumento da quantidade da biomassa da planta; e (iv) melhora no suprimento e ingestão de nutrientes.

Uma das principais vantagens do uso das microalgas como biofertilizantes é a possibilidade de aproveitar a biomassa oriunda dos processos de biorremediação na fertilização de solos. A biomassa destas microalgas é riquíssima em matéria orgânica e substâncias inorgânicas (nitrogênio e fósforo), e sua aplicação como biofertilizantes abre uma perspectiva sobre o conceito de bioeconomia circular e se insere nos ODS estabelecidos pela ONU. Microalgas da espécie C. vulgaris, utilizadas no tratamento de águas residuais de esgoto, foram empregadas como biofertilizantes em cultivo de tomates.59 Estes tomateiros produziram 29 frutos após 3 meses, com pesos médios de até 60 g. Os resultados apresentaram-se promissores, quando comparados com culturas sem adição de fertilizantes (produção de apenas 21 frutos) e culturas com adição de fertilizantes químicos (produção de 33 frutos). Em outro estudo, a biomassa da espécie Chlorella sp. , cultivada em resíduo líquido de aterro sanitário (chorume), produziu biofertilizantes nitrogenados granulares.60 Os resultados demonstraram que a composição química da biomassa obtida, atendeu aos requisitos para a produção do fertilizante nitrogenado, sendo possível granular biofertilizantes contendo até 43% de nitrogênio. Devido à presença de moléculas bioativas na composição de sua biomassa, estudos têm explorado a ação de microalgas como bioestimulantes em cultivares. Os bioestimulantes atuam no processo fisiológico das plantas, incluindo atividades como alongamento de caule, expansão foliar, comprimento e peso da parte aérea, número de folhas, desenvolvimento de frutos, metabolismo secundário e respostas ao estresse biótico e abiótico.61 Nesta perspectiva, a biomassa da microalga D. salina foi empregada no desenvolvimento de um biofilme polimérico como tecnologia alternativa no revestimento de sementes, visando o aumento germinativo e potencialização de compostos bioativos em cultivares de coentro (Corianduum sativum).62 Os resultados obtidos mostraram que o emprego da biomassa no biofilme promoveu não somente o aumento germinativo e o comprimento da raiz, como também o aumento significativo no conteúdo total fenólico e de flavonoides, além de outros metabólitos secundários (Tabela 17 e Figura 16).
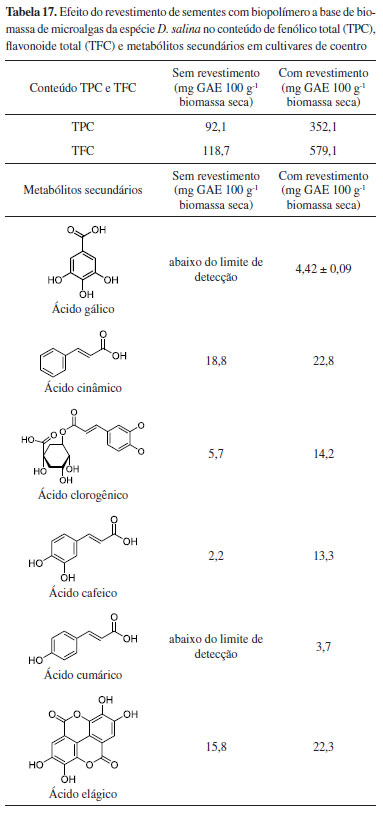
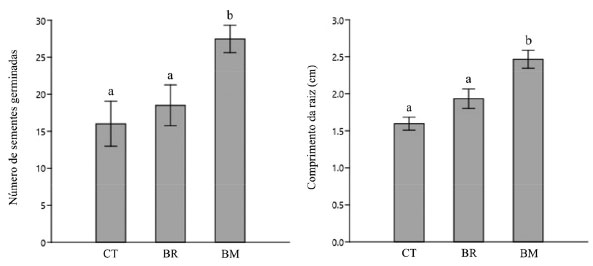 Figura 16. Efeitos do revestimento de sementes com biomassa de microalgas da espécie D. salina nas amostras grupo de controle (CT), branco comparativo (BR) e biopolímero com biomassa de microalgas (BM). Os valores representam as médias da triplicata ± desvio padrão. As diferenças significativas entre as amostras (p < 0,05), medidas pelo teste Kruskal-Wallis e teste de Dunn, são indicadas por letras a e b
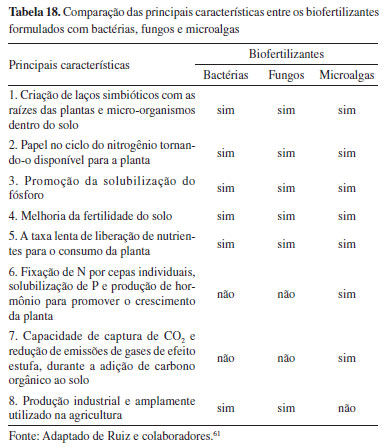
Os valores de TPC são expressos em equivalentes de ácido gálico por 100 g de biomassa seca (mg GAE 100 g-1 biomassa seca) e os valores de TFC são expressos por equivalentes de quercetina por 100 g de biomassa seca (mg QE 100 g-1 biomassa seca). Embora estudos sobre o emprego de microalgas como biofertilizantes tenham apresentado resultados promissores, demonstrando eficiência igual e/ou superior a outros micro-organismos como bactérias e fungos, sua aplicação em escala industrial ainda encontra desafios a serem superados, como alto custo de produção e baixo rendimento na colheita da biomassa (Tabela 18).
Um dos caminhos apontados como estratégia para tais desafios é a aplicação das microalgas para impulsionar a economia circular. Processos integrados surgem como uma alternativa sustentável para obtenção de biofertilizantes, aproveitando a produção de biomassa a partir de processos de biorremediação, como tratamento de efluentes e captura de CO2. Assim, uma vez que as microalgas se mostraram seguras para a formulação de biofertilizantes, capazes de melhorar o rendimento e a qualidade de cultivares em sistemas agrícolas, emerge a necessidade do aprimoramento biotecnológico para implementação desta plataforma em escala industrial.
CONCLUSÕES Devido ao rápido processo de desenvolvimento econômico mundial e alto consumo energético, restringidos pela limitação da disponibilidade de recursos fósseis, existe uma grande demanda pelo desenvolvimento de novas tecnologias renováveis, sustentáveis e menos poluentes. Neste contexto, as microalgas oferecem características únicas para qualificá-las como matéria-prima alternativa para várias aplicações biotecnológicas, incluindo o reaproveitamento de seus resíduos como fonte energética. As microalgas são importantes fontes de metabólitos, como triacilgliceróis, proteínas e compostos de alto valor, como carotenoides e ácidos graxos poliinsaturados de cadeia longa (EPA e DHA). Os triacilgleceróis são matérias-primas essenciais para produção de biocombustíveis, já os carotenoides e os ácidos graxos poliinsaturados podem ser usados como nutracêuticos. Sua capacidade de purificação de águas residuais de diferentes origens, utilizando matéria orgânica como substrato para seu crescimento, aliada à produção de compostos de alto valor agregado, tem mostrado seu potencial como um método econômico e sustentável no tratamento de efluentes. A aplicação das microalgas em processos de Química Verde, como a biocatálise assimétrica de substratos exógenos, demonstra um campo de estudos emergente, com grande potencial para obtenção de compostos quirais, a partir de catalisadores mais eficientes e sustentáveis. Também os avanços sobre o conhecimento genômico de algumas linhagens de microalgas, aliados a ferramentas de engenharia genética, permitem a regulação e produção direcionada de metabólotos específicos, gerando microalgas aprimoradas que levam a processos biotecnológicos otimizados. Por fim, o emprego de microalgas possui um amplo potencial biotecnológico, proporcionando diferentes abordagens tecnológicas integradas a processos químicos, bioquímicos e energéticos para o desenvolvimento sinérgico entre indústria, sociedade e meio ambiente.
AGRADECIMENTOS Os autores agradecem às agências brasileiras CNPq e FAPESB, e ao INCT E&A pelos suportes financeiros. Ao SESI (Serviço Social da Indústria) pelo apoio a F. L. B. M.
REFERÊNCIAS 1. Silva, I. F.; Nascimento, P. H. P.; Lago, R. M.; Ramos, M. N.; Galembeck, F.; Rocha Filho, R. C.; Teixeira, A. P. C.; Quim. Nova 2022, 45, 497. [Crossref] 2. Sutherland, D. L.; McCauley, J.; Labeeuw, L.; Ray, P.; Kuzhiumparambil, U.; Hall, C.; Doblin, M.; Nguyen, L. N.; Ralph, P. J.; Current Research in Environmental Sustainability 2021, 3, 100050. [Crossref] 3. Rizwan, M.; Mujtaba, G.; Memon, S. A.; Lee, K.; Rashid, N.; Renewable Sustainable Energy Rev. 2018, 92, 394. [Crossref] 4. Kokabi, K.; Gorelova, O.; Ismagulova, T.; Itkin, M.; Malitsky, S.; Boussiba, S.; Solovchenko, A.; Goldberg, I. K.; Plant Sci. 2019, 283, 95. [Crossref] 5. Andrade, D. S. Em Microalgas Continentais: Potencialidades e Desafios do Cultivo; Andrade, D. S.; Colozzi Filho, A. , eds.; IAPAR: Londrina, 2014. 6. Kholssi, R.; Ramos, P. V.; Marks, E. A. N.; Montero, O.; Rad, C.; Biocatal. Agric. Biotechnol. 2021, 36, 102114 [Crossref]; Oliveira, D. T.; Costa, A. A. F.; Costa, F. F.; Rocha Filho, G. N.; Nascimento, L. A. S.; Molecules 2020, 25, 2908. [Crossref] 7. Peng, L.; Fu, D.; Chu, H.; Wang, Z.; Qi, H.; Environ. Chem. Lett. 2020, 18, 285 [Crossref]; Pereira, C. M. P.; Hobuss, C. B.; Maciel, J. V.; Ferreira, L. R.; Del Pino, F. B.; Mesko, M. F.; Lopes, E. J.; Colepicolo Neto, P.; Quim. Nova 2012, 35, 2013 [Crossref]; Franco, A. L. C.; Lôbo, I. P.; Cruz, R. S.; Teixeira, C. M. L. L.; Almeida Neto, J. A.; Menezes, R. S.; Quim. Nova 2013, 36, 437. [Crossref] 8. Chai, W. S.; Tan, W. G.; Munawaroh, H. S. H.; Gupta, V. K.; Ho, S. H.; Show, P. L.; Environ. Pollut. 2021, 269, 116236. [Crossref] 9. Levasseur, W.; Perré, P.; Pozzobon, V.; Biotechnol. Adv. 2020, 41, 107545. [Crossref] 10. Posse, F. G.; Figueroa, L. B.; Arias, J. H.; Sánches, D. G.; Molecules 2018, 23, 1265. [Crossref] 11. Braun, J. C. A.; Colla, L. M.; Bioenergy Res. 2023, 16, 289. [Crossref] 12. Yadav, G.; Sen, R.; J. CO2 Util. 2017, 17, 188. [Crossref] 13. Brembu, T.; Muhlroth, A.; Alipanah, L.; Bones, A. M.; Philos. Trans. R. Soc. , B. 2017, 372, 1. [Crossref] 14. Waine, C. K.; Reen, C. S.; Show, P. L.; Jiun, Y. Y.; Chuan, L. T.; Shu, C. J.; J. Taiwan Inst. Chem. Eng. 2018, 91, 332. [Crossref] 15. Chen, X.; Li, Z.; He, N.; Zheng, Y.; Li, H.; Wang, H.; Wang, Y.; Lu, Y.; Li, Q.; Peng, Y.; Biotechnol. Biofuels 2018, 11, 190. [Crossref] 16. Yu, X.; Chen, L.; Zhang, W.; Frontiers in Microbiology 2015, 6, 56. [Crossref] 17. Fabris, M.; Abbriano, R. M.; Pernice, M.; Sutherland, D. L.; Commault, A. S.; Hall, C. C.; Labeeuw, L.; McCauley, J. I.; Kuzhiuparambil, U.; Ray, P.; Kahlke, T.; Ralph, P. J.; Front. Plant Sci. 2020, 11, 279. [Crossref] 18. Chye, J. T. T.; Lau, Y. J.; Yon, J. L. S.; Pan, S.; Danquah, M. K. Em Bioenergy and Biofuels; Konour, O. , ed.; CRC Press: Boca Raton, 2017, cap. 3. [Crossref] 19. Thanh, N. T.; Mostapha, M.; Lam, M. K.; Ishak, S.; Dasan, Y. K.; Lim, J. W.; Tan, I. S.; Lau, S. Y.; Chin, B. L. F.; Hadibarata, T.; Energy Convers. Manage. 2022, 270, 116212. [Crossref] 20. Dall'Oglio, D. F.; Sousa, L. C.; Sousa, S. A. A.; Garcia, M. A. S.; Sousa, E. S.; Lima, S. G.; Costa, P. S.; Guldhe, A.; Bux, F.; Moura, E. M.; Moura, C. V. R.; J. Braz. Chem. Soc. 2019, 30, 633. [Crossref] 21. Wahidin, S.; Idris, A.; Yusof, N. M.; Kamis, N. H. H.; Shaleh, S. R. M.; Energy Convers. Manage. 2018, 171, 1397. [Crossref] 22. Pinho, D. M. M.; Oliveira, R. S.; Santos, V. M. L.; Marques, W. F.; Pinto, A. C.; Rezende, M. J. C.; Suarez, P. A. Z.; J. Braz. Chem. Soc. 2017, 28, 2429. [Crossref] 23. Silva, C. E. F.; Bertucco, A.; Braz. Arch. Biol. Technol. 2019, 62, 1. [Crossref] 24. Torres, G. M. F.; Mahmood, W. M. A. W.; Pittman, J. K.; Theodoropoulos, C.; Biochem. Eng. J. 2020, 153, 107396. [Crossref] 25. Ho, S. H.; Zhang, C.; Tao, F.; Zhang, C.; Chen, W. H.; Trends Biotechnol. 2020, 38, 1023. [Crossref] 26. Chen, Y. C.; Chen, W. H.; Lin, B. J.; Chang, J. S.; Ong, H. C.; Appl. Energy 2016, 181, 110. [Crossref] 27. Vingiani, G. M.; Luca, P.; Ianora, A.; Dobson, A. D. W.; Lauritano. C.; Mar. Drugs 2019, 17, 459. [Crossref] 28. Dias, G.; Hipólito, M.; Santos, F.; Lourega, R.; Mattia, J.; Eichler, P.; Alves, J.; Quim. Nova 2019, 42, 891. [Crossref] 29. Wey, Y. H.; Wen, C. P.; Yuan, H. C.; Ling, L.; J. Taiwan Inst. Chem. Eng. 2017, 74, 1. [Crossref] 30. Wang, L.; Xiao, H.; He, N.; Sun, D.; Duan, S.; Sci. Rep. 2019, 9, 5277. [Crossref] 31. Wang, P.; Wong, Y. S.; Tam, N. F. Y.; J. Appl. Phycol. 2017, 29, 263. [Crossref] 32. Christwardana, M.; Hadiyanto, H.; Motto, S. A.; Sudarno, S.; Haryani, K.; Biomass Bioenergy 2020, 139, 105617. [Crossref] 33. Schüler, L. M.; Schulze, P. S. C.; Pereira, H.; Barreira, L.; León, R.; Varela, J.; Algal Res. 2017, 25, 263. [Crossref] 34. Polishchuk, A.; Valev, D.; Tarvainen, M.; Mishra, S.; Kinnunen, V.; Antal, T.; Yang, B.; Rintala, J.; Tyystärvi, E.; Bioresour. Technol. 2015, 193, 469. [Crossref] 35. Slattery, S. S.; Diamond, A.; Wang, H.; Therrien, J. A.; Lant, J. T.; Jazey, T.; Lee, K.; Klassen, Z.; Penix, I. D.; Karas, B. J.; Edgell, D. R.; ACS Synth. Biol. 2018, 7, 328. [Crossref] 36. Anila, N.; Simon, D. P.; Chandrashekar, A.; Ravishankar, G. A.; Sarada, R.; Photosynth. Res. 2016, 127, 321. [Crossref] 37. Arendt, P.; Pollier, J.; Callewaert, N.; Goossens, A.; Plant J. 2016, 87, 16. [Crossref] 38. Vavitsas, K.; Fabris, M.; Vickers, C. E.; Genes 2018, 9, 520. [Crossref] 39. Wichmann, J.; Baier, T.; Wentnagel, E.; Lauersen, K. J.; Kruse, O.; Metab. Eng. 2018, 45, 211. [Crossref] 40. Lauersen, K. J.; Baier, T.; Wichmann, J.; Wördenweber, R.; Mussgnug, J. H.; Hübner, W.; Huser, T.; Kruse, O.; Metab. Eng. 2016, 38, 331. [Crossref] 41. Lauersen, K. J.; Wichmann, J.; Baier, T.; Kampranis, S. C.; Pateraki, I.; Møller, B. L.; Kruse, O.; Metab. Eng. 2018, 49, 116. [Crossref] 42. Bordón, D. L.; Vázquez, A. M.; Decarlini, M. F.; Demmel, G. I.; Rossi, L. I.; Aimar, M. L.; Biocatal. Biotransform. 2021, 39, 1. [Crossref] 43. Victor, M. M.; Uchôa, J. E.; Riatto, V. B.; Ind. Crops Prod. 2017, 96, 126 [Crossref]; Riatto, V. B.; Victor, M. M.; Sousa, J. F.; Menegola, C.; J. Braz. Chem. Soc. 2019, 30, 149. [Crossref] 44. Yamanaka, R.; Nakamura, K. Em Future Directions in Biocatalysis; Matsuda, T. , ed.; Elsevier: Amsterdam, 2017, cap. 3. [Crossref] 45. Zheng, L.; Zhang, X.; Bai, Y.; Fan, J.; Algal Res. 2018, 35, 432. [Crossref] 46. Schmermund, L.; Jurkas, V.; Özgen, F. F.; Barone, G. D.; Büchsenschütz, H. C.; Winkler, C. K.; Schmidt, S.; Kourist, R.; Kroutil, W.; ACS Catal. 2019, 9, 4115. [Crossref] 47. Chen, G.; Zhang, S.; Hu, Q.; Deng, X.; Yu, F.; Yang, Z.; Hou, Y.; Adv. Mater Res. 2013, 803, 60 [Crossref]; Moutinho, F. : Avaliação do Potencial Biocatalítico da Microalga Dunaliella salina na Redução Assimétrica da Acetofenona; Dissertação de Mestrado, Universidade Federal da Bahia, Bahia, Brasil, 2017. [Link] acessado em Setembro 2023 48. Duda, E. Z.; Głąb, A.; Górak, M.; Ochab, M. K.; Rodak, M. B.; Strub, D.; Śliżewska, A.; Bioorg. Chem. 2019, 93, 102810. [Crossref] 49. Jodlbauer, J.; Rohr, T.; Spadiut, O.; Mihovilovic, M. D.; Rudroff, F.; Trends Biotechnol. 2021, 39, 875. [Crossref] 50. Luo, W.; Deng, X. X.; Gong, Z. W.; Yang, Z. H.; Asia-Pac. J. Chem. Eng. 2016, 11, 533. [Crossref] 51. Tanaka, S.; Kojima, H.; Takeda, S.; Yamanaka, R.; Takemura, T.; Tetrahedron Lett. 2020, 61, 151973. [Crossref] 52. Tanaka, S.; Kojima, H.; Takeda, S.; Yamanaka, R.; Takemura, T.; Tetrahedron Lett. 2021, 77, 153249. [Crossref] 53. Löwe, J.; Siewert, A.; Scholpp, A. C.; Wobbe, L.; Gröger, H.; Sci. Rep. 2018, 8, 10436. [Crossref] 54. Sengupta, A.; Sunder, A. V.; Sohoni, S. V.; Wangikar, P. P.; J. Biothechnol. 2019, 289, 1. [Crossref] 55. Nakamura K.; Yamanaka, R.; Tetrahedron: Asymmetry 2002, 13, 2529. [Crossref] 56. Singh, D.; Thapa, S.; Geat, N.; Mehriya, M. L.; Rajawat, M. V. S. Em Biofertilizers; Rakshit, A.; Meena, V. S.; Parihar, M.; Singh, H. B.; Singh, A. K. , eds.; Woodhead Publishing: Sawston, 2021, cap. 12. [Crossref] 57. Freire, R. C. M.; Aguiar, A. C. M.; Nascimento, M. A.; Cruz, F. S. O.; Mounteer, A. H.; Silva, A. A.; Lopes, R. P.; J. Braz. Chem. Soc. 2023, no prelo [Crossref]; Romão, D. C. F.; Santana Junior, C. C.; Brito, M. R.; Scapin, E.; Pedroza, M.; Rambo, M. C. D.; Rambo, M. K. D.; J. Braz. Chem. Soc. 2022, 33, 938. [Crossref] 58. Cao, T. N. D.; Mukhtar, H.; Le, L. T.; Tran, D. P. H.; Ngo, M. T. T.; Pham, M. D. T.; Nguyen, T. B.; Vo, T. K. Q.; Bui, X. T.; Sci.Total Environ. 2023, 870, 161927. [Crossref] 59. Pooja, K.; Priyanka, V.; Rao, B. C. S.; Raghavender, V.; Energy Nexus 2022, 7, 100122. [Crossref] 60. Slinksienė, R.; Sendzikiene, E.; Mikolaitiene, A.; Makareviciene, V.; Paleckiene, R.; Ragauskaite, D.; Green Chem. Lett. Rev. 2022, 15, 416. [Crossref] 61. Osorio-Reyes, J. G.; Valenzuela-Amaro, H. M.; Pizaña-Aranda, J. J. P.; Ramirez-Gamboa, D.; Meléndez-Sánchez, E. R.; López-Arellanes, M. E.; Castañeda-Antonio, M. D.; Coronado-Apodaca, K. G.; Araújo, R. G.; Sosa-Hernández, J. E.; Melchor-Martínez, E. M.; Iqbal, H. M. N.; Parra-Saldivar, R.; Martinez-Ruiz, M.; Mar. Drugs 2023, 21, 93. [Crossref] 62. Fonseca, Y. B. T.; Almeida, N. M.; Caldas, J. C.; Morais, G. N.; Silva, I. M. J.; Riatto, V. B.; Santos, W. N. L.; Moutinho, F. L. B.; An. Acad. Bras. Cienc. 2022, 94, 1. [Crossref] |
On-line version ISSN 1678-7064 Printed version ISSN 0100-4042
Qu�mica Nova
Publica��es da Sociedade Brasileira de Qu�mica
Caixa Postal: 26037
05513-970 S�o Paulo - SP
Tel/Fax: +55.11.3032.2299/+55.11.3814.3602
Free access